Нейронная сеть, отвечающая за эксплицитные воспоминания - с точки зрения нейрофизиологии
Эксперименты на лабораторных животных, в основном на крысах и обезьянах, позволили ученым воспроизвести подобные наблюдаемым у пациентов Г. М. и Дж. К. симптомы, вызвав у животных поражения медиальных областей височных долей и базальных ядер соответственно. Другие структуры, прежде всего лобные и височные доли, также участвуют в формировании определенных типов эксплицитных воспоминаний.
Теперь рассмотрим отвечающие за эксплицитную и имплицитную память системы по отдельности.
Тяжелый амнестический синдром, обнаруженный у пациентки Г. М. в 1950-х гг., заставил ученых обратить внимание на такую структуру, как гиппокамп, который в то время считали крупной мозговой структурой, и сосредоточиться на поисках его функции. Однако у Г. М. были повреждены и другие области, и первоначальные представления о гиппокампе как о месте обработки эксплицитной информации оказались ошибочными.
P.S. Гиппокамп отвечает за видоспецифичное поведение, ориентирование в пространстве и память; кроме того, он чувствителен к стрессу.
После нескольких десятилетий анатомических и поведенческих исследований, позволивших преодолеть ряд сложностей, к середине 1990-х гг. ученым удалось прийти к единому мнению относительно анатомии эксплицитной памяти. За эксплицитную память в первую очередь отвечают медиальная височная доля, префронтальная кора и тесно связанные с ними структуры.
P.S. Обратившись к литературе, опубликованной до 1995 г., вы можете найти описания механизмов памяти, совершенно отличные от приведенных в данном разделе сайта.
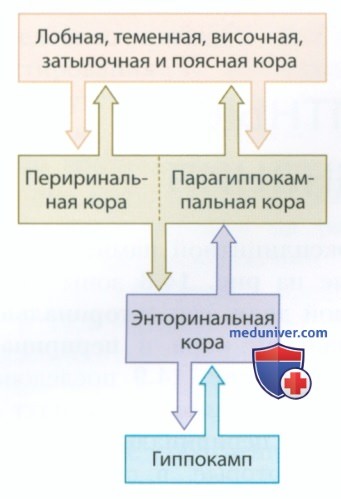
По своему анатомическому строению медиальная область височной доли макаки очень похожа на соответствующую область головного мозга человека. Помимо гиппокампа и миндалины, в формировании эксплицитной памяти участвуют такие изображенные на рис. 1 зоны медиальной области височной доли, как энторинальная кора, парагиппокампальная кора и периринальная кора. Изображенные на рис. 2 последовательно расположенные двусторонние связи идут из основных областей коры в периринальную и парагиппокампальную кору, которые, в свою очередь, проецируются на энторинальную кору, а затем на гиппокамп.
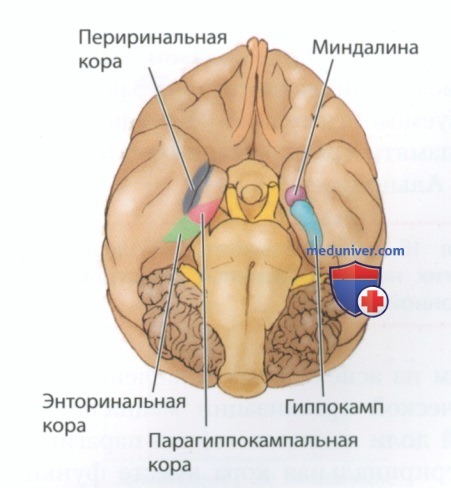
P.S. Периринальная кора (от греч. para — рядом и rhino — нос) лежит рядом с носовой бороздой в нижней части головного мозга.
Обширный корковый вход поступает в периринальную область из вентрального потока обработки зрительной информации, проходящего через височную долю. Таким образом, периринальная область является основным предполагаемым местом локализации зрительно-предметной памяти. Точно так же парагиппокампальная кора получает обширный вход из областей теменной доли, которые, по имеющимся данным, участвуют в формировании зрительно-пространственной памяти — использовании зрительной информации для извлечения воспоминания о пространственном расположении объекта.
Поскольку и периринальная, и парагиппокампальная области проецируются на энторинальную кору, эта область, по-видимому, задействована в обеспечивающих память преимущественно интегративных процессах. Фактически энторинальная кора представляет первую область, где начинают гибнуть нейроны при болезни Альцгеймера, так называемом нейрокогнитивном расстройстве, характеризуемом тяжелыми нарушениями эксплицитной памяти (разд. ниже «Клинические аспекты: Болезнь Альцгеймера»).
Одним из аспектов представленной на рис. 2 анатомической организации медиальных областей височной доли является то, что парагиппокампальная и периринальная кора вместе функционируют как «ворота» в энторинальную кору и гиппокамп. Несмотря на свою популярность, данная точка зрения не учитывает наличие огромного количества связей между префронтальной корой и гиппокампом (более подробную информацию можно найти у Murray et al, 2017). Вскоре мы вернемся к обсуждению роли взаимодействия между префронтальной корой и гиппокампом и их участием в формировании памяти.
а) Гиппокамп и пространственная память. Мы так и не разгадали загадку. Если гиппокамп не является основной отвечающей за эксплицитную память структурой, несмотря на то что на него проецируется энторинальная кора, то какова его функция? Джон О’Киф и Линн Надель (John O’Keefe, Lynn Nadel) были первыми учеными, в 1978 г. выдвинувшими гипотезу о том, что гиппокамп может быть задействован в необходимых для запоминания мест процессах зрительно-пространственной памяти, например в извлечении воспоминания о расположения объекта в пространстве.
Безусловно, и лабораторные животные, и люди с определенными поражениями гиппокампа демонстрируют признаки серьезных нарушений различных форм пространственной памяти. Точно так же обезьяны с поражениями гиппокампа испытывают трудности с запоминанием расположения объектов (зрительно-пространственное научение) — наглядным примером является выполнение заданий, подобных представленному на рис. 3.
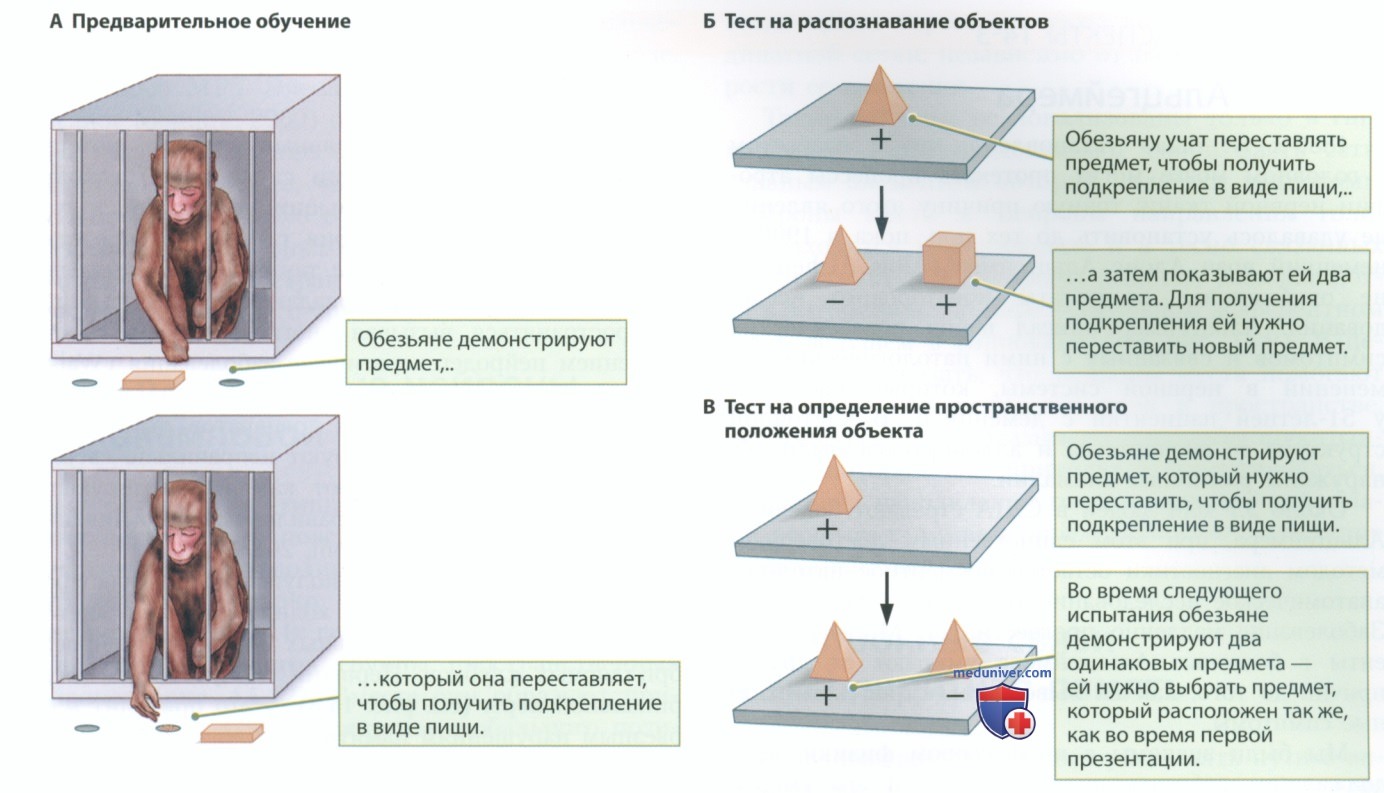
Обезьян учили переставлять предметы, чтобы получить подкрепление в виде пищи (рис. 3, А), а затем давали им одно из двух заданий. В тесте на зрительное распознавание (рис. 3, Б) животное переставляет предмет, чтобы получить подкрепление. После короткой паузы животному показывают два предмета, в том числе один новый предмет. Задача состоит в том, чтобы научиться переставлять новый предмет и получить подкрепление. Данный тест предназначен для оценки эксплицитной зрительной памяти на предметы. Обезьяны с поражениями периринальной коры не справляются с этим заданием.
В тесте на определение положения объекта (рис. 3, В) обезьяне показывают один предмет, который нужно переместить, чтобы получить подкрепление в виде пищи. Затем ей показывают тот же самый предмет, рядом с которым находится второй такой же предмет. Задача состоит в следующем: научиться переставлять предмет таким образом, чтобы он оказался именно в том месте, где находился во время первого предъявления.
Обезьяны с поражениями гиппокампа не способны справиться с отдельными элементами этого задания.
Такие результаты позволяют предположить, что животные с особенно хорошей пространственной памятью должны иметь более крупный гиппокамп, чем животные с менее развитой пространственной памятью. Дэвид Шерри и его коллеги (David Sherry, 1992) решили проверить эту гипотезу на птицах.
Многие виды птиц делают запасы: они собирают семена подсолнечника или другую любимую пищу и прячут, чтобы съесть позднее. Многие птицы способны находить сотни запасенных ими предметов. Чтобы понять, участвует ли в производстве такой деятельности гиппокамп, Шерри и его коллеги измерили размеры гиппокампа у родственных видов птиц — при этом склонность к запасанию пищи демонстрировали представители только одного из двух видов.
Как показано на рис. 4, гиппокамповая формация крупнее у птиц, которые запасают пищу, чем у птиц, которые не делают запасы. В действительности гиппокамп у делающих запасы птиц в два с лишним раза крупнее, чем можно было бы ожидать у птиц с такими размерами мозга и массой тела.
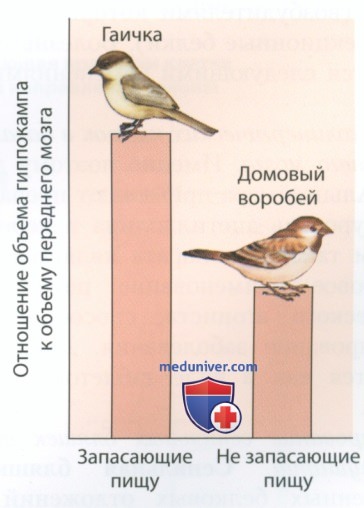
Шерри обнаружил такую же тенденцию, когда сравнил разные виды запасающих пищу грызунов. Кенгуровый прыгун Мерриама — грызун, прячущий запасы на всей территории обитания, — имеет более крупный гиппокамп, чем знамехвостый кенгуровый прыгун, который прячет пищу лишь в собственной норе. По-видимому, и у птиц, и у млекопитающих размеры гиппокампа связаны с когнитивными аспектами двух требующих пространственной ориентации форм деятельности -добывания пищи и запасания пищи.
Одно из предположений, которое мы можем сделать, основываясь на экспериментах Шерри, заключается в том, что у людей, чья работа требуетхороших пространственных способностей, должен быть более крупный гиппокамп. К этой категории можно отнести лондонских таксистов. Чтобы успешно сдать экзамен и получить водительское удостоверение для управления лондонским кэбом, соискатель должен показать, что знает расположение всех улиц в этом огромном древнем городе.
С помощью МРТ Элеонор Магуайр и ее коллеги (Eleanor Maguire, 2000) обнаружили, что у лондонских таксистов задние области гиппокампа были существенно крупнее, чем у испытуемых из контрольной группы. Это открытие позволяет объяснить, почему отдельным людям удается справиться с тестами на пространственную память, которые большинство из нас с треском проваливает.
б) Клинические аспекты: Болезнь Альцгеймера. В 1880-х гг. было установлено, что с возрастом в головном мозге могут протекать процессы атрофии нервной ткани; точную причину этого явления не удавалось установить до тех пор, пока в 1906 г. немецкий врач Алоис Альцгеймер (Alois Alzheimer) не опубликовал результаты новаторского исследования. Альцгеймер описал набор поведенческих симптомов и связанных с ними патологических изменений в нервной системы, которые наблюдал у 51-летней пациентки с деменцией. В клеточной структуре ее неокортекса и аллокортекса были обнаружены различные аномалии.
Около 5,7 млн людей в США страдают болезнью Альцгеймера, при этом единственным достоверным методом диагностики остается посмертное патологоанатомическое исследование тканей головного мозга. Заболевание медленно прогрессирует. Многие пациенты с болезнью Альцгеймера умирают от других причин до того, как их выводят из строя когнитивные симптомы.
Мы были знакомы с профессором физики, продолжавшим работать вплоть до самой его смерти от сердечного приступа в возрасте около 80 лет. Посмертное патологоанатомическое исследование головного мозга профессора выявило значительные, характерные для болезни Альцгеймера изменения. Коллеги профессора списывали его проблемы с памятью на старческий возраст.
Причины болезни Альцгеймера неизвестны. При этом ее часть связывают с генетической предрасположенностью, аномальным уровнем микроэлементов (например, алюминия), иммунными реакциями, медленными вирусными инфекциями и прионными инфекциями (возбудителями которых являются аномальные инфекционные белки). Болезнь Альцгеймера сопровождается следующими изменениями в нервной системе:
1. Гибель холинергических клеток в базальных отделах переднего мозга. Именно поэтому для лечения болезни Альцгеймера применяют препараты, повышающие уровень ацетилхолина в переднем мозге. Примером такого препарата является Экселон® — это торговое наименование ривастигмина, холинергического агониста, способного замедлить прогрессирование заболевания. Данный препарат выпускается как в виде таблеток, так и в виде пластыря.
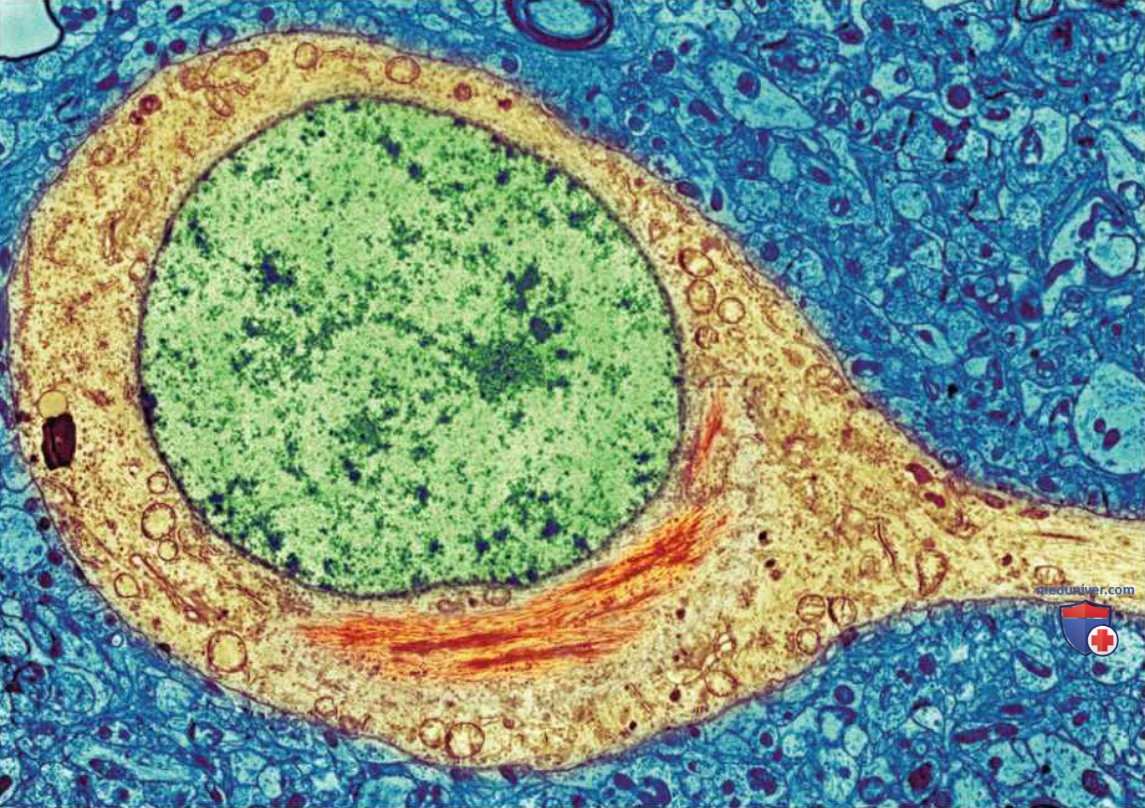
2. Формирование сенильных бляшек в коре больших: полушарий. Сенильная бляшка состоит из гомогенных белковых отложений (амилоид), окруженных фрагментами клеток с признаками дегенеративных изменений. Бляшки распределены в коре неравномерно — они накапливаются прежде всего в связанных с памятью областях височных долей. Формирование сенильных бляшек часто связано с другой аномалией — формированием нейрофибриллярных клубков, которые образуются при агрегации спиральных белковых филаментов в коре больших полушарий и гиппокампе.
Прионная теория гласит, что представленные на рисунке ниже тау-белки с аномальной третичной структурой обладают способностью распространяться, вызывая многие связанные со старением нейродегенеративные заболевания (Walker & Jucker, 2015). Исследователи проводят клинические испытания новых препаратов, которые обнаруживают и нейтрализуют неправильно свернутые белки или действуют как иммунизирующие агенты, препятствуя неправильному сворачиванию белков (Wisniewski & Goni, 2015).
Нейроны коры больших полушарий начинают разрушаться по мере гибели холинергических клеток, а также накопления сенильных бляшек и нейрофибриллярных клубков. Первыми гибнут клетки энторинальной коры (см. рис. 1) — это приводит к серьезным нарушениям памяти.
Выдвинутая неврологами, занимающимися лечением инсультов, спорная теория гласит, что деменция может быть следствием хронических цереброваскулярных нарушений — критически повышенного артериального давления. Повышение артериального давления до критических значений может привести к церебральным микрокровоизлияниям, прежде всего в белом веществе. Образующиеся за годы или даже десятилетия микрокровоизлияния становятся причиной прогрессирующих когнитивных нарушений. В начале могут возникнуть легкие когнитивные нарушения, которые медленно прогрессируют по мере возникновения новых микрокровоизлияний (см. Arvanitakis et al., 2018).
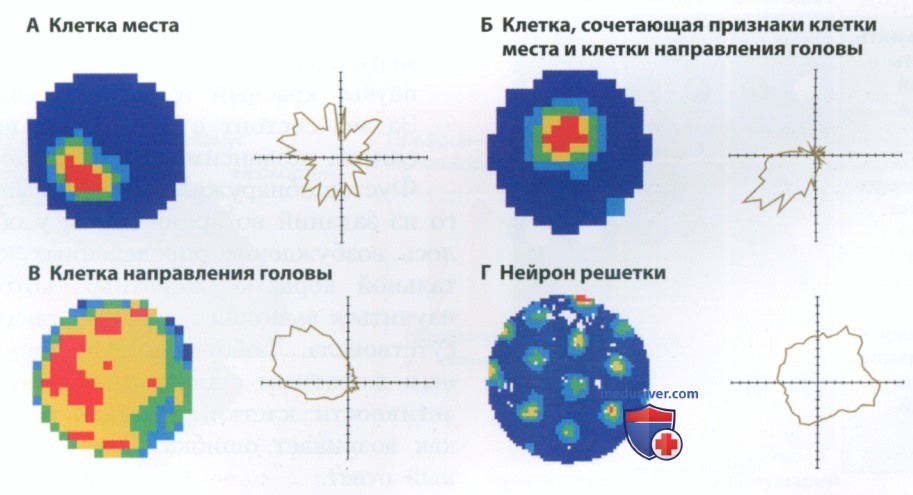
в) Пространственные нейроны гиппокамповой формации. Зная об участии гиппокампа в формировании пространственного поведения, мы можем предположить, что за кодирование пространственной информации отвечают отдельные клетки. Это действительно так. В гиппокампе крыс и мышей были обнаружены три группы пространственных нейронов (рис. 5). Вместе они образуют природный аналог GPS — системы глобального позиционирования. Такие нейроны активно разряжаются, когда животное оказывается в определенной точке пространства.
В 2014 г. Джон О’Киф, Эдвард Мозер и Мэй-Бритт Мозер (John O’Keefe, Edvard Moser, May-Britt Moser) получили за это открытие Нобелевскую премию по физиологии и медицине.
Представленные на рис. 5, А и Б нейроны места возбуждаются в этот момент, когда крыса оказывается в определенном месте, независимо от ориентации ее тела. Нейроны направления головы (рис. 5, В) разряжаются, когда голова крысы ориентирована в определенном направлении. Нейроны решетки (рис. 5, Г) разряжаются, когда крыса пересекает узлы воображаемой координатной сетки, независимо от направления и скорости ее движения.
Такие нейроны не локализованы только в гиппокампе — они представляют собой часть сети, главным элементом которой является гиппокамп. Нейроны места и нейроны направления головы находятся в гиппокампе и в тесно связанных с ним структурах. Нейроны решетки располагаются в энторинальной коре — основном афферентном пути, ведущем в гиппокамп (см. рис. 2). В целом можно сказать, что нейроны места помогают определить положение объектов в пространстве, нейроны решетки показывают масштабы пространства, в котором нам приходится ориентироваться, а нейроны направления головы вместе с нейронами решетки помогают определить наше положение в пространстве.
г) Реципрокные связи и эксплицитная память. Обеспечивающие эксплицитную память пути в височных долях являются реципрокными: неокортекс проецируется на энторинальную кору, которая, в свою очередь, проецируется на неокортекс (см. рис. 2). Реципрокные связи обладают двумя преимуществами:
1. Сигналы, идущие из медиальных областей височной доли обратно в сенсорные области коры, обеспечивают сохранение сенсорного опыта в головном мозге: репрезентация переживания в нервной системе сохраняется дольше, чем фактическое переживание.
2. Путь, ведущий обратно в неокортекс, снабжает его информацией, перерабатываемой в медиальных областях височной доли.
Несмотря на то что мы сконцентрировали свое внимание на медиальных областях височной доли, другие структуры также играют важную роль в формировании эксплицитной памяти. Люди с повреждениями лобных долей не теряют память, как Г. М. или Дж. К. Вместо этого у них появляются проблемы с запоминанием хронологии событий. Представьте себе, будто бы вам показали серию фотографий и попросили запомнить их. Через несколько минут вас спрашивают, узнаете ли вы две фотографии, и просят показать, какую из них вы увидели первой.
Г. М. не запомнил бы фотографии. Люди с поражениями лобных долей вспомнили бы, что видели фотографии, но им было бы трудно вспомнить, какую из них они видели последней. Очевидно, что лобная доля играет менее заметную роль в формировании эксплицитной памяти, чем медиальные области височной доли.
P.S. Системы базальных ядер, которые принимают участие в формировании имплицитной памяти, не осуществляют обратную связь в кору больших полушарий — этим можно объяснить неосознаваемый характер имплицитной памяти.
ЛОБНАЯ ДОЛЯ И КРАТКОВРЕМЕННАЯ ПАМЯТЬ. Все сенсорные системы мозга передают информацию в лобные доли, как и медиальные области височной доли. Эта информация не используется непосредственно для сенсорного анализа, то есть у нее должно быть другое назначение. В общем и целом лобная доля участвует в формировании многих форм кратковременной памяти.
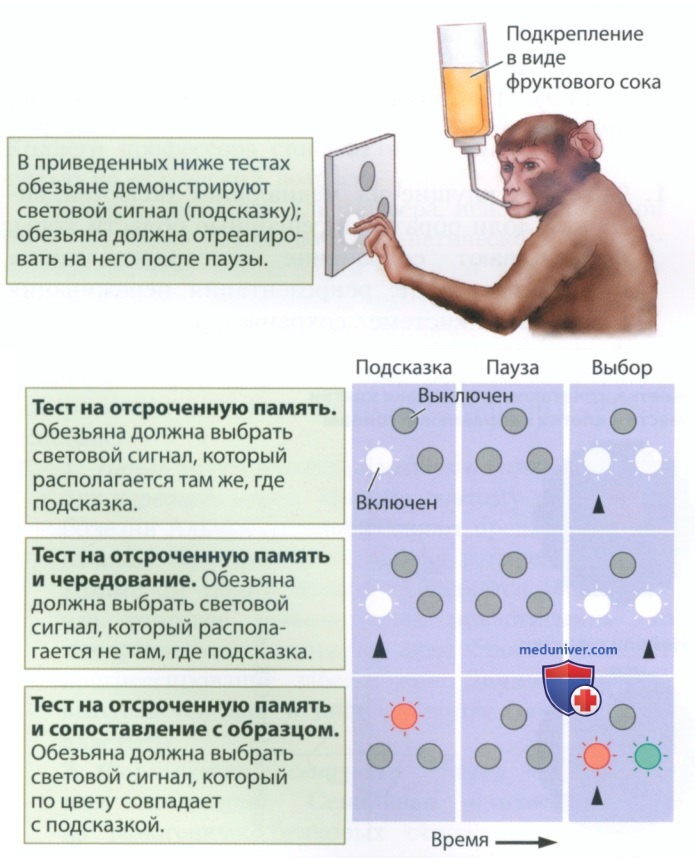
Хоакин Фустер (Joaquin Faster; например, Faster et al., 2000) измерял потенциалы отдельных клеток лобной коры во время выполнения заданий на кратковременную память. Например, если обезьянам показывают объект, который они должны запомнить на короткое время, прежде чем им позволят дать ответ, нейроны префронтальной коры продолжают оставаться в возбужденном состоянии на протяжении всей паузы. Давайте рассмотрим тесты, представленные на рис. 6:
• дизайн всех экспериментов предполагает, что обезьяне демонстрируют световой сигнал (подсказку); после паузы обезьяна должна отреагировать на него, чтобы получить подкрепление;
• в тесте на отсроченную память обезьяне демонстрируют два световых сигнала — ей необходимо выбрать тот сигнал, который находится там, где была подсказка;
• в тесте на отсроченную память и чередование обезьяне снова демонстрируют два световых сигнала — теперь ей нужно выбрать тот сигнал, который находится не там, где была подсказка;
• в тесте на отсроченную память и сопоставление с образцом обезьяне демонстрируют, к примеру, красный световой сигнал, а затем, после паузы, красный и зеленый световые сигналы. Задача состоит в том, чтобы выбрать красный сигнал, независимо от его местоположения.
Фустер обнаружил, что при выполнении каждого из заданий во время паузы у обезьян наблюдалось возбуждение определенных клеток префронтальной коры. У животных, которые не смогли научиться выполнять задание, такая активность отсутствовала. Любопытно, что допущенные обученным животным ошибки отражаются на клеточной активности: клетки перестают реагировать до того, как возникает ошибка. Они «забывают» правильный ответ.
НЕЙРОННАЯ СЕТЬ ЭКСПЛИЦИТНОЙ ПАМЯТИ. У людей, длительное время злоупотребляющих алкоголем, может возникнуть расстройство эксплицитной памяти, называемое синдромом Корсакова. В некоторых случаях тяжелые нарушения эксплицитной памяти распространяются и на имплицитную память. Синдром Корсакова развивается из-за нехватки тиамина (витамина B1), приводящего к гибели нейронов в медиальных областях промежуточного мозга (разделяет головной мозг и ствол мозга), в том числе в медиальном таламусе и мамиллярных телах гипоталамуса. У 80% пациентов с синдромом Корсакова наблюдается атрофия (утрата клеток) лобных долей.
По-видимому, тяжесть нарушений памяти связана с тем, что поражения затрагивают не только передний мозг, но и структуры ствола мозга (разд. ниже «Клинические аспекты: Синдром Корсакова»).
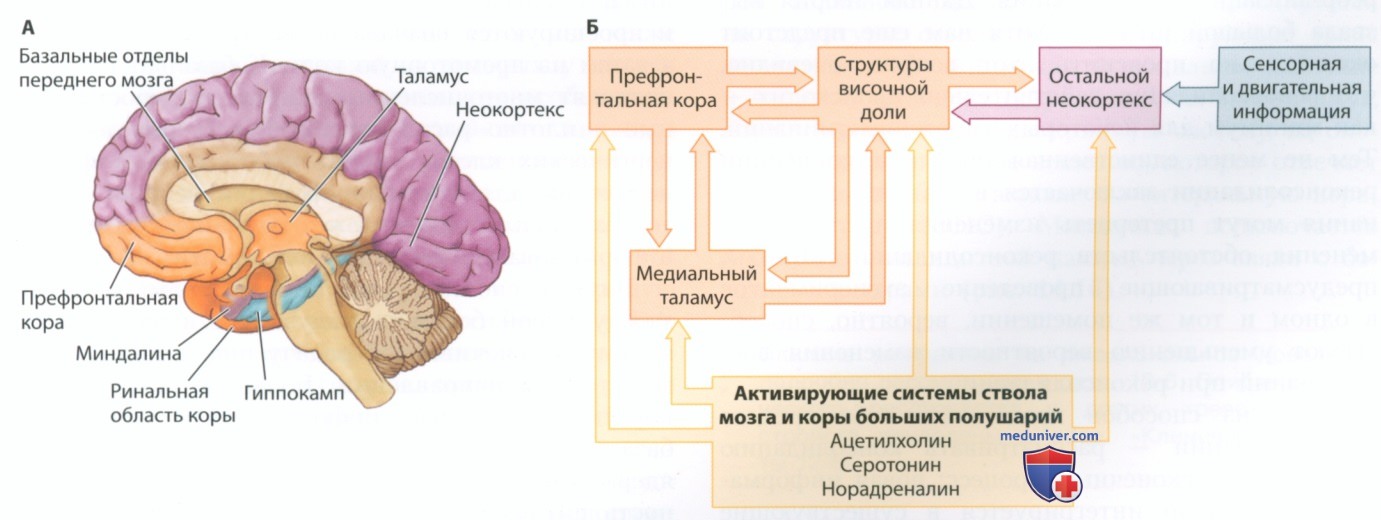
Мортимер Мишкин и его коллеги (Mortimer Mishkin; Mishkin, 1982; Murray, 2000) смоделировали нейронную сеть эксплицитной памяти, основываясь на данных исследований людей и лабораторных животных с повреждениями височных и лобных долей. На рис. 7 представлена модифицированная версия созданной Мишкиным модели. Анатомически (рис. 7, А) она задействует не только лобную и височную доли, но также медиальную область таламуса, которая связана с развитием синдрома Корсакова, и системы активации базальных отделов переднего мозга, связанные с развитием болезни Альцгеймера. На рис. 7, Б представлена схема информационных потоков:
• сенсорные и моторные области неокортекса проецируются на медиальные области височной доли, которые, в свою очередь, проецируются на медиальную область таламуса и префронтальную кору;
• ученые предполагают, что структуры базальных отделов переднего мозга играют некую роль в поддержании определенного уровня активации других структур переднего мозга, давая им возможность обрабатывать информацию;
• считается, что структуры височной доли и префронтальной коры играют ведущую роль в формировании долговременной эксплицитной памяти;
• префронтальная кора играет главную роль в обеспечении кратковременного хранения эксплицитных воспоминаний, а также в запоминании хронологии эксплицитных событий.
д) Клинические аспекты: Синдром Корсакова. Хронический алкоголизм, особенно в сочетании с неправильным питанием, нарушает память. Когда 62-летнего Джо Р. госпитализировали, его семья жаловалась, что его память стала отвратительной. Джо Р. обладал средним уровнем интеллекта, у него отсутствовали очевидные сенсорные или двигательные нарушения.
Когда его спрашивали, чем он занимался накануне вечером, Джо Р. обычно отвечал: «Ходил в «Легион» выпить несколько стаканчиков пива с парнями». Хотя в действительности Джо Р. находился в больнице, такой ответ можно было считать вполне разумным — посещение «Легиона» было тем, чем Джо Р. занимался почти каждый вечер на протяжении последних 30 лет.
Джо Р. точно не знал, чем зарабатывает на жизнь, но считал себя мясником. На самом деле он водил грузовик, принадлежавший местной службе доставки. Однако мясником был его сын, то есть в истории Джо Р. все-таки фигурировали какие-то связанные с его жизнью факты.
Память Джо Р. на недавние события была не лучше. В одной ситуации мы попросили его запомнить нашу встречу, а затем вышли из помещения. Вернувшись спустя 2-3 минуты, мы обнаружили, что он не помнит ни встречу, ни то, что проходил предложенные нами психологические тесты.
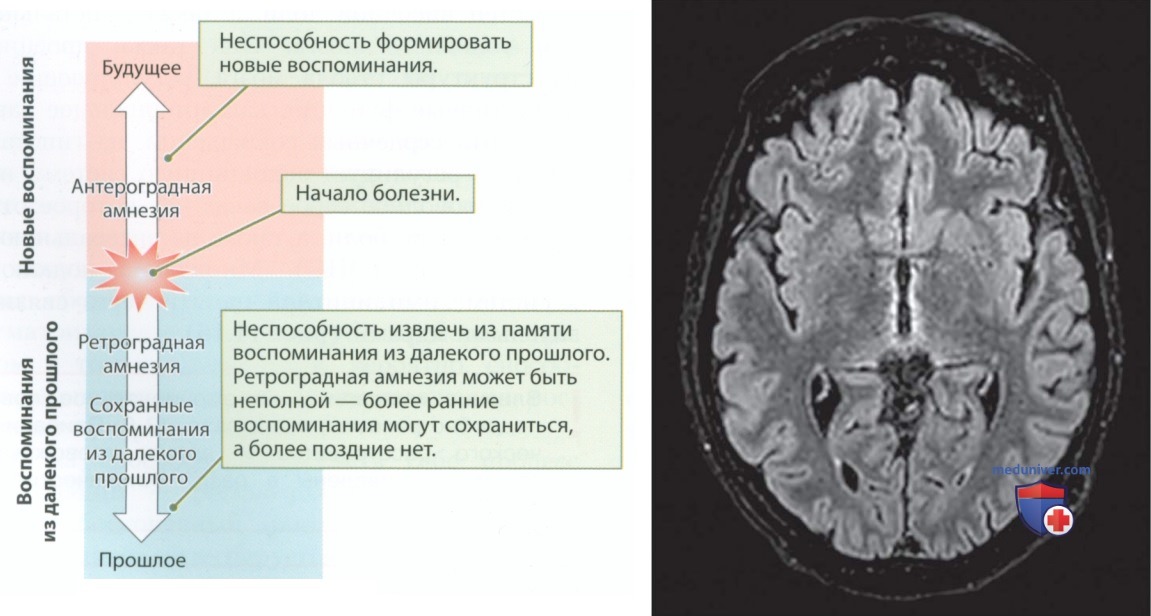
У Джо Р. был синдром Корсакова. Сергей Корсаков был русским врачом, который в 1880-х гг. обратил внимание на синдром, развивающийся при хроническом алкоголизме. Наиболее очевидным симптомом являются тяжелые нарушения памяти, в том числе отсутствие воспоминаний о событиях, происходивших до начала амнезии (ретроградная амнезия), и потеря способности запоминать события, происходящие после возникновения нарушения памяти (антероградная амнезия).
Уникальной особенностью пациентов с синдромом Корсакова является то, что они склонны сочинять истории о своем прошлом вместо того, чтобы признать, что ничего не помнят. Как и истории Джо Р., такие истории обычно правдоподобны, потому что основаны на реальном опыте.
Интересно, что пациенты с синдромом Корсакова не отдают себе отчета в том, что у них есть нарушения памяти. Когда им указывают на проблемы с памятью, они, как правило, демонстрируют безразличие. Кроме того, такие пациенты обычно равнодушны к тому, что происходит вокруг.
Причиной развития синдрома Корсакова является нехватка тиамина (витамина Bt), возникающая из-за плохого питания или продолжительного употребления больших доз алкоголя. (Кроме «нескольких стаканчиков пива с парнями», Джо Р. на протяжении многих лет ежедневно выпивал по бутылке рома в 26 унций.) Нехватка тиамина приводит к гибели клеток в области срединной линии промежуточного мозга, прежде всего в медиальных областях таламуса и мамиллярных телах гипоталамуса.
У большинства пациентов с синдромом Корсакова возникает атрофия коры больших полушарий, особенно в лобных долях. После появления симптомов синдрома Корсакова, которые могут возникнуть внезапно, прогноз неблагоприятный. Только у 20% пациентов состояние улучшается после года применения обогащенного витамином рациона. За несколько лет состояние Джо Р. не улучшилось, и он провел последние 15 лет своей жизни в больнице.
- Читать далее "Консолидация эксплицитных воспоминаний - с точки зрения нейрофизиологии"
Редактор: Искандер Милевски. Дата публикации: 2.11.2023
- Два типа памяти - с точки зрения нейрофизиологии
- Какова природа различий между эксплицитной и имплицитной памятью? - с точки зрения нейрофизиологии
- Каковы особенности личных воспоминаний? - с точки зрения нейрофизиологии
- Диссоциация систем памяти - с точки зрения нейрофизиологии
- Нейронная сеть, отвечающая за эксплицитные воспоминания - с точки зрения нейрофизиологии
- Консолидация эксплицитных воспоминаний - с точки зрения нейрофизиологии
- Нейронная сеть, отвечающая за имплицитные воспоминания - с точки зрения нейрофизиологии
- Нейронная сеть, отвечающая за эмоциональные (аффективные) воспоминания - с точки зрения нейрофизиологии
- Эволюция систем памяти - с точки зрения нейрофизиологии
- Долговременная потенциация как основа нейропластичности - с точки зрения нейрофизиологии