Вторичная соматосенсорная кора (SII) - с точки зрения нейрофизиологии
Вторичная соматосенсорная кора (SII) не только получает соматосенсорную информацию из SI, но и принимает сигналы из зрительной и слуховой коры. В экспериментах на обезьянах электростимуляция SII вызывает движения, похожие на те, которые были вызваны электростимуляцией моторной коры.
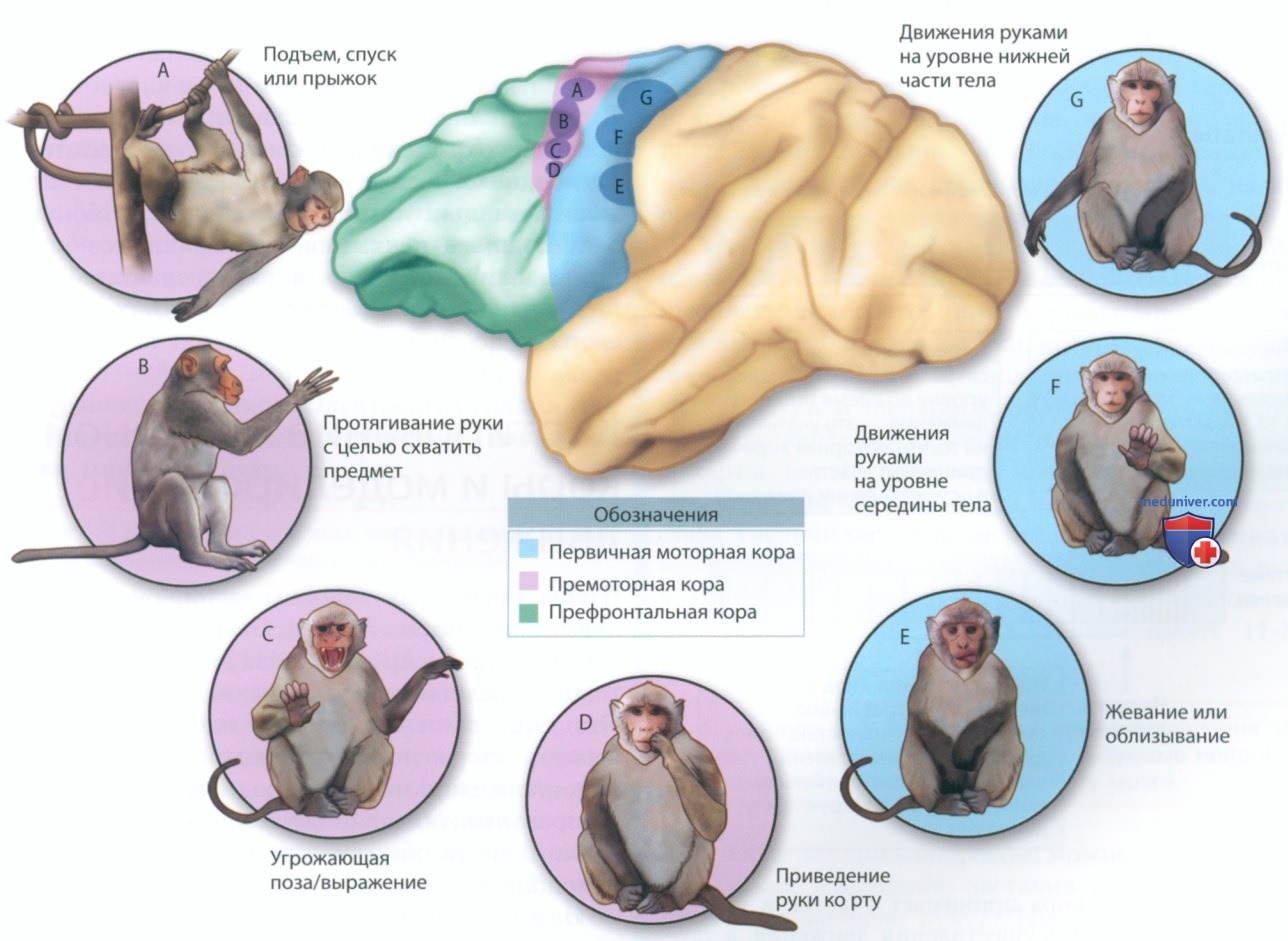
Стимуляция дорсальных областей SII вызывает движения всем телом, такие как прыжки и карабкание вверх; стимуляция вентральных областей SII заставляет протягивать конечности к различным точкам на теле или ко рту. Такая топографическая организация движений в SII не только идентична топографической организации движений в моторной коре — анатомическое строение SII указывает на наличие связей между функциональными зонами области SII и моторной коры.
Наличие связей между соматосенсорной корой и моторной корой наводит на мысль о том, что движение берет свое начало в соматосенсорной коре. Каков же на самом деле вклад соматосенсорной коры в производство движения? Вероятно, ответ состоит в том, что зрительные и слуховые области коры определяют пространственное расположение тех объектов вокруг нас, с которыми мы могли бы взаимодействовать.
Эта информация передается в соматосенсорную кору теменной доли, определяющей, какая часть тела будет взаимодействовать с расположенными в этом месте объектами. Затем эта информация передается в моторную кору, которая отвечает за выполнение движения.
Пример такой организации движения представлен на рис. ниже. На рисунке изображены три пути, или канала, — дорсальный путь, обеспечивающий протягивание руки, вентральный, отвечающий за хватание, и расположенный между двумя предыдущими путь, обеспечивающий приведение руки ко рту. Пути, ответственные за протягивание руки и хватание объекта, образуют связи со зрительной корой, а путь, обеспечивающий приведение руки ко рту, не образует.
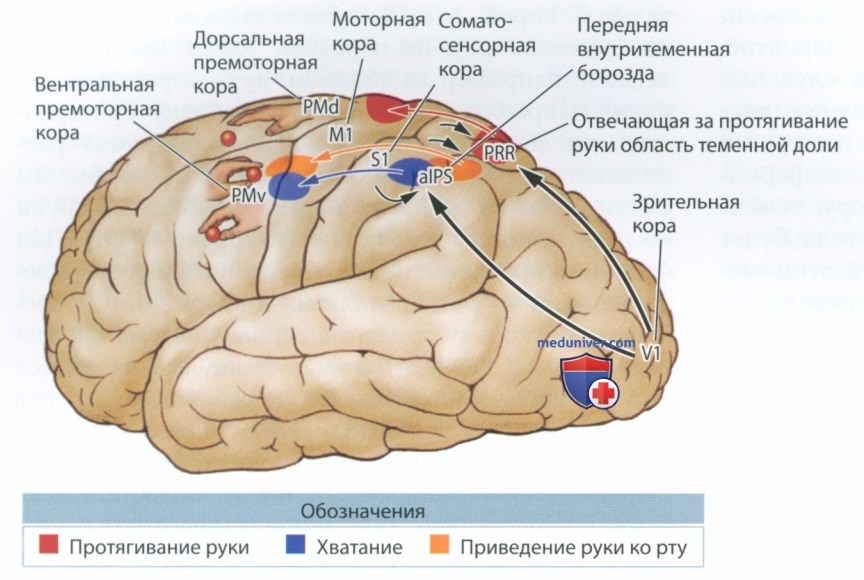
Когда мы тянемся за едой, например за яблоком, путь «протягивания руки» направляет нашу руку в ту точку пространства, где находится яблоко, путь «хватания» располагает наши пальцы таким образом, чтобы мы могли схватить яблоко, а путь «приведения руки ко рту» помогает нам поднести яблоко ко рту. Для функционирования пути, обеспечивающего приведение руки ко рту, зрительная информация не требуется, поскольку соматосенсорная система определяет, что мы держим яблоко, знает, где находится рот, и может организовать перемещение яблока ко рту (Karl et aL, 2018).
Этот пример наглядно демонстрирует, что для производства движения используется ограниченный набор подобных изображенным на рис. ниже элементарных движений. Такие элементарные движения можно сравнить со словами, переставляя которые можно получить разные предложения.
а) Исследование: Щекотка. Щекотка, как и ее последствия, знакома всем. Это ощущение представляет собой удивительное сочетание приятных и неприятных ощущений. Существует два вида щекотки: книсмезис — ощущение легких прикосновений, и гаргалезис — ощущение более грубых ритмических прикосновений.
Щекотку ощущают не только люди, но и другие приматы, а также кошки, крысы и, возможно, большинство других млекопитающих. Игровое поведение у крыс ассоциировано с вокализациями. Щекотание участков тела, с которыми обычно играют крысы, заставляет крыс издавать сигнал с частотой 50 кГц (LaFollette et al., 2017).
Щекотка приносит удовольствие — и люди, и животные любят, когда их щекочут. Им также нравится смотреть, как щекочут других. Использование робота и методов нейровизуализации позволило Саре Блэкмор и ее коллегам (Sarah Blakemore, 1998) найти объяснение тому, почему человек не может пощекотать самого себя.
Эксперимент Блэкмор предполагал воздействие на ладони испытуемых идентичных тактильных стимулов двух типов. В первом случае стимул был ожидаемым, а во втором случае робот предъявлял стимул с непрогнозируемой задержкой. Только неожиданный стимул вызывал ощущение щекотки. Таким образом, ощущение щекотки обусловлено не самой стимуляцией, а ее непредсказуемостью. Именно поэтому человек не может пощекотать самого себя — он знает, что делает.

Проанализировав самостоятельные отчеты испытуемых, Уиндт и ее коллеги (Windt, 2015) обнаружили, что в осознанных снах стирается грань между ощущениями, создаваемыми самим испытуемым, и ощущениями, возникающими в результате действий другого человека: людям снится, как они щекочут сами себя.
Одной из интересных особенностей щекотки является вызываемый ею характерный смех. Такой смех можно идентифицировать на сонограмме (графическое представление звука); люди в состоянии отличить вызываемый щекоткой смех от других видов смеха. В отношениях между матерью и ребенком смех, вызываемый щекоткой, может способствовать укреплению взаимной привязанности (Ishijima & Negayama, 2017).
Узнав о том, что все высшие обезьяны смеются, когда их щекочут, Росс и его коллеги (Ross, 2010) решили сравнить вызываемый щекоткой смех у разных высших обезьян и обнаружили, что смех человека больше похож на смех шимпанзе, чем на смех горилл и других высших обезьян. Это значит, что восприимчивость к щекотке и смех мы унаследовали от общего предка высших обезьян.
- Читать далее "Последствия повреждения соматосенсорной коры - с точки зрения нейрофизиологии"
Редактор: Искандер Милевски. Дата публикации: 10.10.2023
- Вторичная соматосенсорная кора (SII) - с точки зрения нейрофизиологии
- Последствия повреждения соматосенсорной коры - с точки зрения нейрофизиологии
- Соматосенсорная кора и сложные движения - с точки зрения нейрофизиологии
- Резюме по регуляции нервной системой движения и реакции на стимуляцию
- Определение причин эмоционального и мотивационного поведения - с точки зрения нейрофизиологии
- Обонятельная чувствительность - с точки зрения нейрофизиологии
- Вкусовая чувствительность - с точки зрения нейрофизиологии
- Нейроанатомическая основа мотивационного поведения - с точки зрения нейрофизиологии
- Управление пищевым поведением - с точки зрения нейрофизиологии
- Регуляция потребления жидкости - с точки зрения нейрофизиологии