Стволовые лимфоидные клетки - общий лимфоидный предшественник
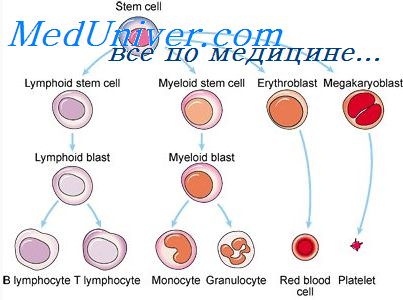
Многообразие типов кроветворных клеток возникает из общей предшественницы — стволовой гемопоэтической клетки (СГК), способной дифференцироваться в направлении любых ростков гемопоэза.
По мере пролиферации и дифференцировки стволовых гемопоэтических клеток (СГК) их по-липотентность утрачивается, так что потомки этих клеток становятся способными к дифференцировке по ограниченному числу гемопоэтических линий, впоследствии лишь по одной из них. Клетки-предшественницы и СГК экспрессируют на мембране антиген CD34, который отсутствует на зрелых клетках.
В костном мозге человека присутствует немногочисленная субпопуляция стволовых гемопоэтических клеток (СГК, экспрессирующих на мембране CD34, CD90 (Thyl), но не имеющих маркеров гемопоэтических линий (Lin). На этих клетках отсутствуют миелоидно-моноцитарные антигены (CD13, CD33, CD14, CD15), специфические маркеры эритроидной (гликофорин А) и мегакариоцитарной (CD41, CD42, CD61) линий, В-лимфоидные антигены (CD19, CD20), маркеры Т-клеток и их субпопуляций (CD7, CD5, CD2, CD3, CD4, CD8), а также маркеры ЕК-клеток (CD16, CD56).
Стволовая гемопоэтическая клетка (СГК) человека (CD34+Lin-) при длительном культивировании (4—6 нед) способны давать начало миелоидным (CD33+), В-лимфоидным и Т-лимфоидным клеткам. Мультипотентными, т. е. способными к дифференцировке по всем гемопоэтическим линиям, являются клетки CD34+ Lin-, CD38", HLA-DR+ в пределах СГК.
Лимфоидная дифференцировка стволовых клеток изучена недостаточно. Схематично взаимоотношения стволовой гемопоэтической клетки и общего лимфоидного предшественника (ОЛП) представлены на рисунке. Из СГК возникает общий лимфоидный предшественник. ОЛП впервые обнаруживаются в онтогенезе в аортогонадно-мезонефральной области плода человека с 5-й недели внутриутробного развития. Эти клетки дают начало лишь В-, Т- и ЕК-клеткам.
Общие лимфоидные клетки-предшественницы человека экспрессируют CD34, CD 10, CD38, HLA-DR и CD45RA, но не имеют на мембране CD90, CD7, CD117 и различных линейно-ассоциированных маркеров.
На ранних этапах дифференцировки лимфоидных предшественников решающее значение для их созревания имеет экспрессия специфичных для лимфоидных клеток генов. Экспрессия этих генов регулируется рядом транскрипционных факторов (ТФ), необходимых для развития Т- или В-клеток либо свойственных всем трем лимфоидным линиям. Отсутствие ТФ приводит к остановке дифференцировки Т-, В- или NK-клеток на ранних стадиях созревания, что проявляется в виде той или иной формы иммунодефицита.
Следствием возникающих в результате отсутствия ТФ иммунодефицитных состояний является более частое возникновение опухолей. Так, дефицит Ets-1 приводит к снижению количества NK-клеток и развитию NK-чувствительных опухолей. Нередко возникают опухоли из клеток поврежденной лимфоид-ной линии. При мутациях в генах Ikaros или Е2А развиваются Т-клеточные лимфомы.
В случаях отсутствия Рах5 (другое название «специфичный для В-клеток активаторный белок» — BSAP; от англ. B-cell-specific activator protein) дифференцировка В-лимфоцитов протекает только до стадии про-В, не происходит реаранжировки генов д-цепей иммуноглобулина на этапе V-DJ. Про-В-клетки, не имеющие Рах5, экспрессируют гены других гемопоэтических линий: миелопероксидазу, М-КСФ, GATA-1 (эритроидные), перфорин (NK), рТа (Т) — и могут дифференцироваться в макрофаги, остеокласты, дендритные клетки, гранулоциты и NK-клетки.
Восстановление активности Рах5 в про-В-клетках подавляет экспрессию антигенов неродственных линий. Этот пример важен для понимания возможных механизмов экспрессии антигенов разных гемопоэтических линий на уровне одной клетки, что является достаточно распространенным явлением при острых лейкозах.
Наиболее характерная особенность созревания Т-и В-лимфоцитов — сборка функциональных генов рецепторов для антигена (ГРА) - Ig (BCR) и TCR из отдельных фрагментов (V, D, J или V, J), разобщенных в геноме зародышевых клеток. Процесс формирования ГРА протекает при обязательном участии белков, кодируемых генами, активирующими рекомбинацию (RAG-1 и RAG-2; от англ. recombination-activating genes), ферментов терминальной дезоксирибонуклеотидилтрансферазы (TdT) и ДНК-зависимой протеинкиназы (ДНК-ПК).
Дефицит белков RAG-1, RAG-2, TdT, ДНК-ПК приводит к нарушениям развития Т- и В-лимфоцитов на ранних стадиях, что отражается на их количестве или качественном составе и может повысить частоту лимфом.
- Читать "Онтогенез В-лимфоцитов - дифференциация"
Оглавление темы "Гемопоэз":- Механизмы миграции стволовых кроветворных клеток (СКК)
- Мезенхимная стволовая клетка - свойства, функции
- Эмбриональная стволовая клетка (ЭСК) - свойства, функции
- Пластичность стволовых клеток - возможности дифференцировки
- Врожденный и адаптивный иммунитет - механизмы
- Стволовые лимфоидные клетки - общий лимфоидный предшественник
- Онтогенез В-лимфоцитов - дифференциация
- Перестройка иммунноглобулинов при В-лимфоцитопоэзе
- Антигены и рецепторы В-лимфоцитов
- Созревание В-лимфоцитов в костном мозге