Мембранный потенциал покоя кардиомиоцитов (КМЦ) сердца (фаза 4 потенциала действия)
Внутриклеточный потенциал во время электрического покоя в течение диастолы колеблется от -50 до -95 мВ в зависимости от типа клетки, поэтому заряд внутри клетки на 50-95 мВ более отрицателен, чем снаружи, что обусловлено распределением таких ионов, как К+, Na+ и Cl-.
Поскольку в КМЦ в состоянии покоя имеется избыток открытых K+-каналов, трансмембранный потенциал (в фазе 4) близок к Еk. Выходящий ток калия через открытые К+-каналы, выпрямляющие входящий ток IKI, в нормальных условиях в значительной мере зависит от мембранного потенциала покоя предсердных и желудочковых КМЦ, а также клеток Пуркинье.
Отличие мембранного потенциала покоя от Еk обусловлено движением одновалентных ионов с равновесным потенциалом большим, чем Еk, например током Cl- через активированные хлоридные каналы, такие как ICi.camp, ICl.Ca и IЕCl.swellКальций напрямую не влияет на мембранный потенциал покоя, но изменения концентрации свободного внутриклеточного кальция могут отразиться на других показателях мембранной проводимости.
Например, повышение нагрузки Са2+ в СР может обусловить появление спонтанных кальциевых волн, которые, в свою очередь, активируют Са2+-зависимую хлоридную проводимость ICl.Ca и, следовательно, приводят к возникновению спонтанных транзиторных входящих токов и сопутствующей мембранной деполяризации. Повышение [Са2+], также может стимулировать Na+/Ca2+-обмен (INa/Ca). Этот белок меняет 3 иона Na+ на 1 ион Са2+; направление зависит от концентрации натрия и кальция по обе стороны мембраны и разницы трансмембранного потенциала.
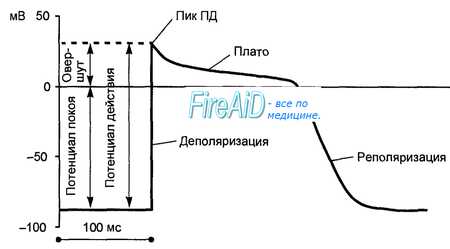
Во время мембранного потенциала покоя и во время спонтанного высвобождения Са2+ в СР этот обменник будет генерировать итоговый ток Na+, возможно вызывающий временную мембранную деполяризацию. Также было показано, что [Са2+], активирует IKI в КМЦ и таким образом косвенно влияет на кардиальный мембранный потенциал покоя. Поскольку Na+/K+-нacoc перекачивает Na+ из клетки против его электрохимического градиента и одновременно отправляет К+ в клетку также против его электрохимического градиента, концентрация внутриклеточного К+ остается высокой, а концентрация внутриклеточного Na+ — низкой.
Этот насос, энергетический ресурс которого обеспечивается ферментом Na+/К+-АТФазой, гидролизирующей АТФ для высвобождения энергии, связан с мембраной. Для его работы необходимы и Na+, и К+; он может перемещать 3 иона Na+ из клетки и 2 иона К+ — внутрь клетки. Следовательно, насос может быть электрогенным, а итог его работы — направленный наружу поток положительных зарядов.
Скорость работы Na+/K+-насоса для увеличения тех же ионных градиентов может повышаться подобно ЧСС, поскольку клетка при каждой деполяризации накапливает небольшое количество ионов Na+ и теряет небольшое количество ионов К+.
Гликозид-индуцированное ингибирование Na+/К+-АТФазы повышает сократимость КМЦ за счет увеличения концентрации внутриклеточного Na+, который, в свою очередь, уменьшает вытеснение Са2+ путем Na+/Са2+-обмена, что в итоге и повышает сократимость клеток.

Видео физиология фаз потенциала действия кардиомиоцитов
- Читать "Фаза возбуждения кардиомиоцитов (КМЦ) сердца (фаза 0 потенциала действия)"
Оглавление темы "Фазы потенциала действия кардиомиоцитов сердца":- Строение щелевого соединения вставочных дисков сердца и его функции
- Фазы потенциала действия кардиомиоцитов (КМЦ) сердца
- Мембранный потенциал покоя кардиомиоцитов (КМЦ) сердца (фаза 4 потенциала действия)
- Фаза возбуждения кардиомиоцитов (КМЦ) сердца (фаза 0 потенциала действия)
- Фаза ранней быстрой реполяризации кардиомиоцитов (КМЦ) сердца (фаза 1 потенциала действия)
- Плато потенциала действия кардиомиоцитов (КМЦ) сердца (фаза 2 ПД)
- Фаза конечной быстрой реполяризации кардиомиоцитов (КМЦ) сердца (фаза 3 ПД)
- Фаза диастолической деполяризации кардиомиоцитов (КМЦ) сердца (фаза 4 ПД)
- Механизмы автоматизма синусового узла сердца
- Потеря мембранного потенциала клеткой как причина аритмий