Белок 4.2 клеточной мембраны. Паллидин
Белок 4.2 составляет около 5% всех цитоскелетных белков и представлен более 200000 копиями на клетку. Главной функцией паллидина является регуляция формы и механических свойств эритроцитарной мембраны посредством модификации взаимосвязи анкирина с цитоплазматическим доменом белка 3. Паллидин, связываясь со спектрином, способствует его взаимодействию с анкирином и таким образом участвует в прикреплении мембранного скелета к трансмембранным белкам [(Davies and Lux, 1989)]. Связываясь с цитоплазматическим доменом АТБ, паллидин ограничивает его материальную и вращательную подвижность.
Увеличение количества белка 4.2, связанного с АТБ, приводит к изменению транспорта анионов через клеточную мембрану. Обнаружено, что обработка мембран солевыми растворами, содержащими Mg2+, Ca2+, Cu2+, приводит к усиленному связыванию паллидина с цитоплазматическим сегментом анионного транспортера. По-видимому, посредством такого взаимодействия белок 4.2 регулирует проницаемость мембраны не только для анионов, но и для двухвалентных катионов.
Белок 4.2 в мембране эритроцита существует в виде олигомера сферической формы с Mw=72kD и диаметром 8-15 nm. Существует мнение, что in vivo белок может образовывать и тетрамерные формы с Mw=185kD.
Паллидин имеет несколько сайтов связывания на цитоплазматическом домене белка 3. Потенциально связывающие участки для белка 4.2 на анионном транспортере не полностью заняты в нормальной клетке [266]. Кроме этого, паллидин взаимодействует со спектрином, анкирином и белком 4.1 и может ингибировать реакцию, катализируемую трансглютаминазой, в результате которой образуются сшивки между молекулами цитоскелетных белков.
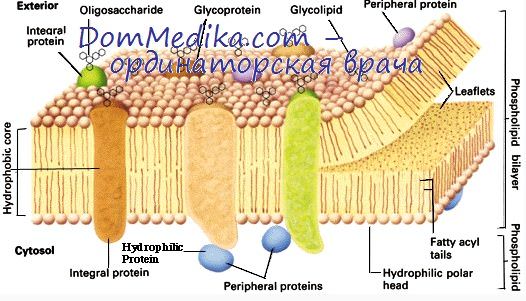
Белок 4.2 может фосфорилироваться цАМФ-зависимыми протеинкиназами. Реакция активируется накоплением ДФГ и подавляется ионами Са2+. Цитоплазматический домен белка 3 ингибирует связывание анкирина и паллидина со спектрин-актиновой сетью.
Korsgren с соавт. (1990) клонировали и секвенировали ДНК белка 4.2 из ретикулоцитов. Белок состоит из 691 аминокислот, кодируемых 2073 парами нуклеотидов. Аминокислотная последовательность белка 4.2 гомологична с 2-мя близко родственными кальций-зависимыми белками -печеночной трансглютаминазой и альфа-субъединицей XIII фактора гемокоагуляции. Белок 4.1 имеет аминокислотную замену, которая нивелирует его трансглютаминазную активность в эритроцитах.
Korsgren и Cohen (1991) показали, что ген белка 4.2 имеет протяженность около 20kb и состоит из 13 экзонов. Ретикулоциты содержат 2 различных по размеру варианта мРНК белка 4.1- главный (меньшего размера) образуется альтернативным сплайсингом. Sung с соавт. (1991) картировали ген белка 4.2 на 15q15-q21. Najfeld (1992) уточнил локализацию этого гена на 15q 15.
- Читать далее "Белок 4.9 клеточной мембраны. Дематин"
Оглавление темы "Строение мембран клеток":1. Ген белка 4.1. Локусы хромосом отвечающие за белок 4.1
2. Белок 4.2 клеточной мембраны. Паллидин
3. Белок 4.9 клеточной мембраны. Дематин
4. Актин клеточной мембраны. Структура и кодирование актина мембран
5. Белок 7 клеточной мембраны. Тропомиозин мембран клеток
6. Тропомодулин клеточной мембраны. Белок 8 мембран клеток
7. Миозин клеточной мембраны. Цилиндрин мембран клеток
8. Белок р55 клеточной мембраны. Белок полосы 3 мембран клеток
9. Функции белка полосы 3. Кодирование белка полосы 3
10. Белок полосы 4.5 мембраны клеток. Структура и кодирование белка полосы 4.5