Трансляция аденовирусов. Этапы трансляции аденовирусов
Размер аденовирусного генома указывает на то, что он способен кодировать около 23 полипептидов со средним мол. весом 50 000 дальтон.
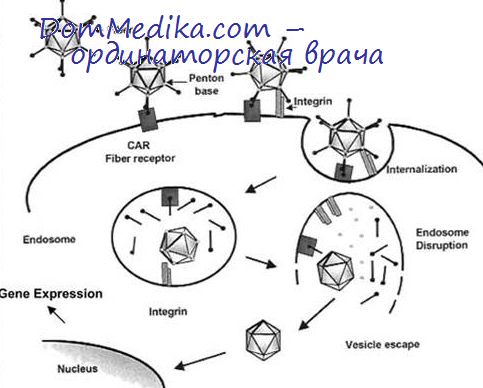
Структурные белки. Вирион содержит несколько структурных белков, молекулярные веса которых лежат в пределах от 22 000 до 120 000. Главные белки — это белки гексона (120 000), основания пентона (70 000), нити (62 000) и сердцевины (46 000 и 22 000). При аденовирусной инфекции эти структурные белки («растворимые антигены») образуются в значительных количествах; время их появления зависит от множественности заражения.
Белки синтезируются в таком избытке, что на поздних стадиях инфекции накапливаются в ядре в виде белковых кристаллов (Годмен и др., 1960).
Все структурные антигены — поздние продукты генома, и их синтез зависит от синтеза вирусной ДНК. Антигены нити и гексона появляются приблизительно через 12—16 ч после заражения, антиген основания пентона — двумя часами позже. Количество инфекционного вируса быстро нарастает параллельно с образованием антигена основания пентона, поскольку с появлением последнего в клетке имеются все компоненты, необходимые для сборки полных вирионов. Свободные пентоны, определяемые методом прямой гемагглютинации, появляются позднее, чем антиген основания пентона.
По предположению Рассела и др. (1971), образовавшиеся антигены основания пентона не включаются в свободные пентоны, а идут преимущественно на сборку вирионов. Большая часть структурных полипептидов образуется ие в результате посттрансляционного расщепления, а синтезируется независимо; белки образуются с различными скоростями в молярных отношениях, не соответствующих их молярным отношениям в вирионе (Уайт и др., 1969).
Например, полипептиды основания пентона и нити образуются в значительном избытке по сравнению с их содержанием в вирионе, а полипептиды сердцевины синтезируются в минимальных количествах (Хорвиц и др., 1969), что предполагает существование некоего механизма, регулирующего транскрипцию и (или) трансляцию мРНК различных видов.
Высокоочищенные вирионы аденовирусов содержат эндонуклеазную активность (Бёрлингхэм и др., 1971). Эта активность появляется в клетках с продуктивным типом инфекции приблизительно в то же время, что и капсидные белки (Бёрлингхэм и Дёрфлер, 1972).
Неструктурные белки. При заражении клеток аденовирусами торможение синтеза белка клетки-хозяина несколько задерживается, поэтому обнаружить кодируемые вирусом неструктурные белки довольно трудно. Многие исследователи показали, что вскоре после заражения синтезируется антиген, иммунологически отличный от капсидных антигенов. Т-антиген выявляется методом диффузии в геле или в реакции связывания комплемента при использовании сывороток хомячков, имеющих индуцированные вирусом опухоли (Бёрмэн и Роу, 1965).
Его биосинтез не зависит от синтеза вирусной ДНК (Гилиэд и Гинс-берг, 1968), т. е. он определяется ранней мРНК.
Используя специфические антисыворотки, приготовленные против очищенных антигенов, Расселу и др. (1967) удалось проследить за образованием антигенов в клетках, зараженных аденовирусом типа 5. Используя кроличью антисыворотку, приготовленную против экстракта клеток, зараженных аденовирусом типа 5 в присутствии ингибитора синтеза ДНК, они выявили другой ранний антиген — Р-антиген. Этот антиген появляется раньше капсидных антигенов; он впервые обнаруживается приблизительно через 5—7 ч после заражения и достигает максимального уровня к 12 ч. В некоторых отношениях Р-антиген сходен с Т-антигеном; это относительно термолабильный ранний антиген, его синтез не зависит от синтеза вирусной ДНК. Функции Т- и Р-антигенов не установлены; по предположению Рассела и др. (1971), Р-антиген является внутренним вирусным белком.
Рассел и Скехел (1972) попытались определить ранние неструктурные полипептиды, пометив зараженные клетки 35S-мeтионином с высокой удельной активностью и проанализировав полипептиды методом электрофореза в геле с последующей радиоавтографией. Через 10 ч после заражения аденовирусом типа 5 они обнаружили 5 неструктурных «специфичных для зараженной клетки» полипептидов (ICSP 1—5). По-видимому, эти полипептиды закодированы в вирусном геноме. ICSP-3, который синтезируется в присутствии цитозинарабинозида и представляет собой основной компонент Р-антигена, определяется уже через 2,5 ч после заражения; другие появляются к 10—13 ч.
Эти полипептиды не синтезируются одновременно с капсидными полипептидами, причем ICSP-1, ICSP-2 и ICSP-3, подобно богатым аргинином полипептидам сердцевины, не синтезируются на поздних сроках инфекции. Рассел и Скехел (1972) подсчитали, что если все «специфичные для зараженной клетки» полипептиды — независимые продукты вирусных генов, то около 15% вирусных полипептидов должно быть отнесено к ранним белкам, что согласуется с данными Грина и др. (см. выше) по определению ранней транскрипции генома. До сих пор в литературе нет четких данных о том, что антисыворотка к Р-антигену реагирует с антигенами, содержащими сердцевинный полипептид 2, хотя существуют косвенные доказательства тождественности сердцевинного полипептида 2 и «позднего» компонента Р-антигена. В настоящее время нет убедительных данных об обнаружении ферментов, индуцированных аденовирусом на ранних сроках инфекции, и биологическая роль ранних «специфичных для зараженной клетки» полипептидов неизвестна.
- Читать далее "Репликация аденовирусной ДНК. Сборка и выход аденовируса из клетки"
Оглавление темы "Паповавирусы. Аденовирусы":1. Цикл размножения паповавирусов. Транскрипция паповавирусов
2. Трансляция паповавирусов. Репликация ДНК паповавирусов
3. Интеграция клеточного и вирусного геномов. Интегративность паповавирусов
4. Сборка и выход паповавирусов из клетки. Метаболизм зараженных паповавирусами клеток
5. Цикл размножения аденовирусов. Начальные стадии аденовирусной инфекции
6. Транскрипция аденовирусов. Механизмы транскрипции аденовирусов
7. Трансляция аденовирусов. Этапы трансляции аденовирусов
8. Репликация аденовирусной ДНК. Сборка и выход аденовируса из клетки
9. Изменение метаболизма инфицированного аденовирусом клетки. Цикл размножения герпесвирусов
10. Начальные стадии герпесвирусной инфекции. Транскрипция герпесвирусов