Начальные стадии герпесвирусной инфекции. Транскрипция герпесвирусов
Существуют противоречивые мнения об относительной важности цитоза и слияния мембран. Хаммелер и др. (1969) предполагают, что может иметь место проникновение обоих типов, но в случае вирионов, имеющих внешнюю липопротеидную оболочку, более важным способом, возможно, является пиноцитоз (виропексис).
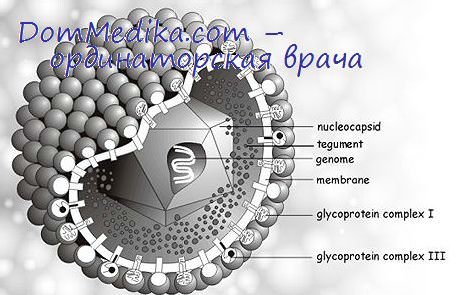
Дискуссия о роли оболочки в инфекционном процессе нашла подробное отражение в работе Дарлингтона и Мооса (1969). Позднее было показано (Эбодили и др., 1970), что захватываться клетками в процессе виропексиса и инициировать инфекцию могут как частицы с оболочкой, так и частицы, оболочка которых удалена в результате химической обработки. Благодаря оболочке, возможно, облегчается адсорбция на клетках и увеличивается эффективность образования бляшек, но ее присутствие не является обязательным условием инфекционности.
К сожалению, пока ничего не известно о том, каким начальным изменениям подвергаются частицы герпесвирусов при попадании внутрь клеток.
Транскрипция герпесвирусов
Хотя герпесвирусы довольно большие и имеют сложную структуру, их вирионы в отличие от вирионов поксвирусов, по-видимому, не несут собственной транскриптазы. Вирус-специфическая РНК образуется первоначально в ядре, возможно с участием клеточной РНК-полимеразы, как это происходит у аденовирусов. Последний довод подкрепляется сообщением (Лэндоу и Райхайнер, 1969), которое, правда, требует подтверждения, об инфекционности ДНК ВПГ.
Вирус-специфическая РНК обнаруживается в ядре в виде молекул с коэффициентами седиментации от 40 до 60S (Вагнер и Ройзман, 1969; Вагнер, 1972). После десятиминутного лаг-периода молекулы РНК, которые гибридизуются с вирусной ДНК (Ройзман и др., 1970) и имеют коэффициенты седиментации в пределах 4—30S (преимущественно 10—20S), появляются в цитоплазме.
Методом конкурентной гибридизации показано, что вирусная ядерная РНК с более высоким молекулярным весом имеет последовательности, общие с цитоплазматической вирусной РНК (Вагнер, 1972); возможно, цитоплазматическая РНК образуется в результате расщепления высокомолекулярных транскриптов, обнаруживаемых в ядре. Герпесвирус-специфические РНК могут быть ранними или поздними, т.е. синтезироваться до или после начала синтеза вирусной ДНК, причем те и другие РНК присутствуют в ядре в виде молекул большего размера, чем в цитоплазме. В опытах по гибридизации немеченой РНК из клеток, зараженных ВПГ, с меченой очищенной вирусной ДНК (Ройзман и Френкель, 1972) выявлено существование двух классов ранней вирусной РНК.
Как на ранних сроках (2 ч после заражения, до синтеза вирусной ДНК), так и на поздних (8 ч после заражения) авторы обнаружили РНК двух отличающихся по количеству молекул классов. С помощью конкурентной гибридизации было показано, что последовательности ранних РНК, относящихся к классу, содержащему большее количество молекул, присутствуют в поздних РНК такого же класса. Авторы предположили, что РНК этих классов определяет синтез некоторых структурных белков вириона, однако данные, подтверждающие это предположение, носят предварительный характер.
В зараженных вирусом псевдобешенства клетках, предварительно обработанных циклогексимидом, образуется вирус-специфическая РНК с коэффициентом седиментации около 26S. Если нет большого избытка вирусной ДНК, то в опытах по ДНК/РНК-гибридизации гибридизуется лишь небольшая доля РНК, что дает основание предположить, что РНК образуется в результате повторяющейся транскрипции небольшого участка вирусного генома (Ракусанова и др., 1971).
Опыты по конкурентной гибридизации и сравнение кривых насыщения вирусной ДНК препаратами РНК, синтезированной в зараженных клетках в присутствии или в отсутствие циклогексимида, привели авторов к заключению, что РНК, синтезирующуюся перед началом синтеза ДНК в зараженных вирусом псевдобешенства клетках (ранняя мРНК), можно разделить на два класса; для одного из них не требуется, а для другого требуется синтез белка после заражения. РНК этих двух классов, возможно, аналогичны «предранним» и «задерживаемым ранним» РНК фага Т4 (Солсер и др., 1970).
В серии работ Субак-Шарпа с соавторами (Субак-Шарп и Хей, 1965; Субак-Шарп и др., 1966а) сообщалось, что вирус простого герпеса может кодировать определенные виды тРНК. Хотя авторы позднее отказались от выводов, сделанных в этих работах (Бэл и др., 1971), но они стимулировали исследования подобного рода для других вирусов животных (см. разделы о поксвирусах и о вирусе саркомы Рауса).
- Вернуться в оглавление раздела "Вирусология"
Оглавление темы "Паповавирусы. Аденовирусы":1. Цикл размножения паповавирусов. Транскрипция паповавирусов
2. Трансляция паповавирусов. Репликация ДНК паповавирусов
3. Интеграция клеточного и вирусного геномов. Интегративность паповавирусов
4. Сборка и выход паповавирусов из клетки. Метаболизм зараженных паповавирусами клеток
5. Цикл размножения аденовирусов. Начальные стадии аденовирусной инфекции
6. Транскрипция аденовирусов. Механизмы транскрипции аденовирусов
7. Трансляция аденовирусов. Этапы трансляции аденовирусов
8. Репликация аденовирусной ДНК. Сборка и выход аденовируса из клетки
9. Изменение метаболизма инфицированного аденовирусом клетки. Цикл размножения герпесвирусов
10. Начальные стадии герпесвирусной инфекции. Транскрипция герпесвирусов