Влияние внутриклеточных факторов на репликацию ВИЧ
Активация Т-клеток и дифференцировка моноцитов могут влиять на уровень продукции вируса. Эти наблюдения подчеркивают роль внутриклеточных факторов в репликативном цикле ВИЧ. Несмотря на то, что длинный концевой повтор (LTR) ВИЧ сам может выступать в роли промотора транскрипции с вирусных генов, ранняя транскрипция мРНК, по-видимому, зависит преимущественно от связывания с LTR клеточных факторов транскрипции, таких как ядерный фактор каппа В (NF-kB), NFAT, АР-1, SP-1 и Tat-связывающие белки.
Таким образом, внутриклеточные факторы могут определять относительное преобладание вирусных регуляторных белков в инфицированной клетке. Анализ микроэрреев с кДНК клеточных генов на материале из CD4+-T-лимфоцитов на стадии острой инфекции, показал, что отличия от неинфицированных клеток касаются генов, ответственных за проведение сигналов, за трафик молекул внутри клетки и генов факторов транскрипции.
Дальнейшие свидетельства участия внутриклеточных факторов в репликации вируса были получены в биологических исследованиях, которые показали различия в репликации одних и тех же изолятов ВИЧ в различных линиях клеток и в мононуклеарных клетках периферической крови, полученных от разных доноров.
Также были описаны различия в чувствительности к инфицированию у макрофагов, выделенных из различных тканей организма. Связывание вируса и его проникновение в клетку может протекать одинаково, но процессы, происходящие после проникновения вируса, могут различаться, так что титры продуцируемых дочерних вирусов варьируют в широком диапазоне. Наиболее ярко такие различия проявляются при изучении ВИЧ-инфекции в мононуклеарных клетках из периферической крови in vitro.
Различные сопутствующие вирусные и внутриклеточные факторы могут влиять на распространение вируса и патогенез заболевания в организме хозяина.
Внутриклеточные блокировки способны нарушать ранние и поздние стадии обратной транскрипции, образование преинтеграционного комплекса и транспорт его в ядро клетки. Например, при аресте клеточного деления Т-лимфоцитов периферической крови вирусная инфекция протекает в абортивной форме, останавливаясь на стадии перед интеграцией провируса ДНК в геном клетки, что преимущественно связано с блокировкой проникновения преинтеграционного комплекса в ядро.
В неделящихся макрофагах или эпителиальных клетках может происходить продукция дочерних вирусов, поскольку в этих клетках PIC способен проникать в интерфазное ядро через ядерную мембрану. CD4+-клетки становятся чувствительными к инфекции после того, как они входят в стадию G1b клеточного цикла. Активация Т-лимфоцита через CD3 (то есть через TCR) сама по себе приводит к переходу клетки в стадию G1a, что создает условия только для незавершенной обратной транскрипции ВИЧ-1.
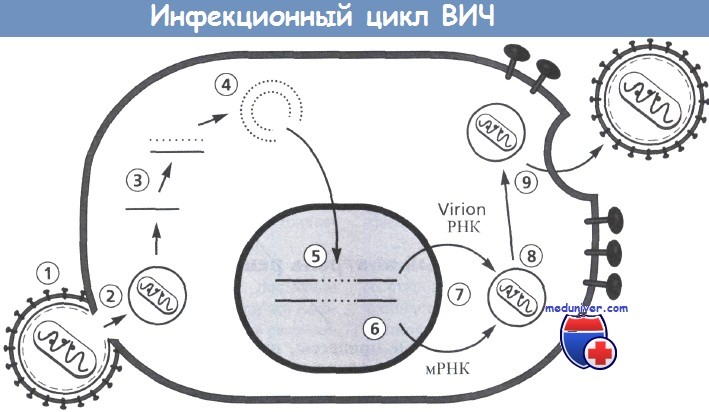
Включает следующие этапы: 1 - прикрепление; 2 - декапсидация; 3 - обратная транскрипция;
4 - образование кольцевых форм; 5 - интеграция; 6 - транскрипция; 7 - трансляция;
8 - сборка вирусной сердцевины; 9 - окончательная сборка и отпочкование вирионов.
На этапах от 3 до 5 некоторые белки вирусной сердцевины связываются с вирусным геномом (РНК, ДНК).
Двухцепочечные циркулярные формы ДНК могут быть связаны ковалентно и нековалентно.
Последние представляют собой формы, которые интегрируются в хромосому клетки. Противовирусная терапия может быть направлена на каждый этап и потенциально способна нарушать репликацию и распространение вируса.
Рисунок предоставлен Н. Kessler.
Совместная стимуляция с участием молекул CD3 (TCR) и CD28 (существенный ко-рецептор TCR) обеспечивает протекание завершенного процесса вирусной транскрипции (2285). В астроцитах уменьшение трансляции вирусных белков, по-видимому, происходит под действием высоких концентраций протеинкиназы, связывающей двухцепочечные РНК, что в свою очередь является результатом низкой экспрессии антагониста этой протеинкиназы - белка, связывающегося с областью молекулы РНК, отвечающей за трансактивацию трансляции (TAR - transactivating response region) (TRBP) (3336).
В пермиссивных активированных Т-клетках интеграция и репликация ВИЧ происходят в течение 24 часов заражения. В макрофагах процесс протекает аналогично, но производство дочерних вирусов может занимать от 36 до 48 часов. Последние данные, возможно, свидетельствуют о более медленном метаболизме предшественников нуклеотидов в неделящихся клетках. Однако при активации макрофагов макрофаг-колониестимулирующим фактором (M-CSF) репликация вируса в этих клетках становится быстрой и высокоэффективной.
Такие различия в репликации ВИЧ исследуют также на моделях введения трансфекции молекулярных клонов кДНК ВИЧ-1 в клетки. В этом случае опять наблюдается вариабельность результатов, зависящая от типа клеток. Обычно репликация ВИЧ в клетках человека происходит с максимальной интенсивностью по сравнению с репликацией в клетках других видов животных, особенно мышей (2534). Аналогичные наблюдения были сделаны при изучении вирусных частиц со смешанным фенотипом (псевдовирионов), которые позволяют вводить геном ВИЧ в гетерологичные клетки с помощью оболочки другого вируса.
После обратной транскрипции с генома их собственной РНК соответствующие вирусы реплицировались в клетках с различной интенсивностью. Внутриклеточный контроль вирусной репликации также наблюдают и в случае других рет-ровирусов. Полученные данные указывают либо на существование клеточных ингибиторов размножения вирусов, либо на относительный недостаток клеточных продуктов, необходимых для репликации вируса.
Некоторые исследования, проведенные на гибридных клетках человека и животных, указывают на доминантность пермиссивного состояния для репликации вируса. Таким образом, по-видимому, отсутствие определенного клеточного белка может ограничивать размножение вируса. Эксперименты на гибридных клетках «человек-хомяк» и «человек-мышь» выявили, что фактор пермиссивности, усиливающий активность Tat, кодируется геном в 12-й хромосоме человека. Рассматривают несколько кандидатных клеточных белков, влияющих на активность Tat. Идентифицирован белок с мол. весом 83 кДа, связывающийся с TAR петлей РНК.
Другие эксперименты показали, что устойчивость клетки к ВИЧ-инфекции также может иметь доминантный характер, и был описан фактор, выделенный из мышиных клеток, подавляющий ген rev. Эти ингибиторы Tat- и Rev-активности в мышиных клетках действуют независимо друг от друга, и их можно рассматривать как потенциальные мишени для разработки методов противовирусной терапии. Открытие противовирусного действия молекулы TRIM5a также имеет отношение к этим данным.
- Читать "Особенности репликации ВИЧ в Т-клетках"
Оглавление темы "Репликация ВИЧ":- Этапы репликации ВИЧ внутри клетки
- Влияние внутриклеточных факторов на репликацию ВИЧ
- Особенности репликации ВИЧ в Т-клетках
- Особенности репликации ВИЧ в моноцитах и макрофагах
- Белок APOBEC3G как механизм резистентности к ВИЧ
- Аналоги белка APOBEC3G у животных
- Природные функции белков APOBEC
- Белок TRIM5a как механизм резистентности к ВИЧ
- Влияние цитокинов на репликацию ВИЧ
- Влияние белков ВИЧ на клетку