Трансляция ортомиксовирусов. Репликация РНК ортомиксовирусов
Вирусные структурные белки синтезируются на цитоплазматических полисомах, но быстро мигрируют в различные участки клетки, причем белок NP накапливается в ядре, а белок НА — на мембранах гладкой эндоплазматической сети (Уайт и др., 1970). Белок М также передвигается в эти мембраны (Лазаровиц и др., 1971; Компанс, 1973).
Главный неструктурный белок NS появляется в ядрышке через 5 мин после синтеза в цитоплазме (Тейлор и др., 1969, 1970; Лазаровиц и др., 1971); кроме того, его обнаруживают в ассоциации с рибосомами (Понс, 1972; Компанс, 1972). Возможно, эти два события взаимосвязаны, так как рибосомы собираются в ядрышке; можно предположить, что белок NS заставляет рибосомы транслировать преимущественно вирусную, а не клеточную мРНК.
Другая возможность заключается в том, что этот белок, синтезирующийся в наибольшем среди вирус-специфических макромолекул количестве, представляет собой субъединицу вирусной РНК-полимеразы (Тейлор и др., 1970). Белок NS почти наверняка является основной субъединицей типоопецифического неструктурного антигена, ранее обнаруженного в ядрышке методом иммунофлуоресценции (Диммок, 1969). Позднее Скехел (1972) сообщил о втором неструктурном белке (мол. вес 11000), выявленном в зараженных клетках с помощью электрофореза в геле.
Клетка, зараженная вирусом гриппа, — хорошая система для изучения факторов, влияющих на внутриклеточное передвижение белков. Было показано, что транспорт в ядро полипептидов NS и NP не требует продолжительного синтеза РНК или белка, а включение в мембраны белка НА — процесс, для которого необходимо поступление энергии (Уайт и др., 1970).
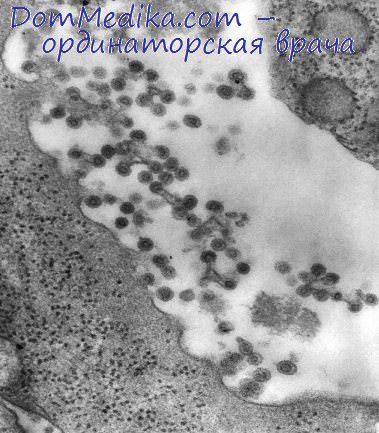
Следует отметить, что белок НА всегда связан с мембранами; он синтезируется на прикрепленных к мембранам рибосомах и в течение нескольких минут перемещается через гладкую эндоплазматическую сеть в плазматическую мембрану (Компанс, 1973; Стенли и др., 1973). Используя различные ингибирующие сахара, которые блокируют завершение образования вирусных гликопротеидов, не влияя на синтез других вирусных белков (Ганди и др., 1972), удалось показать, что почти сразу же после синтеза белка НА в его состав включается глюкозамин, а затем присоединяется фукоза (Стенли и др., 1973).
Гликопротеид НА иногда ферментативно расщепляется на два более мелких гликопротеида, НА1 и НА2, которые остаются соединенными дисульфидными связями (Лазаровиц и др., 1971; Стенли и Хэслем, 1971). Посттрансляционное расщепление является скорее случайным, чем обязательным, так как оно выражено в различной степени в различных комбинациях вирус—клетка, причем некоторые системы дают инфекционные, гемагглютинирующие вирионы, в которых белок НА совсем не расщеплен (Стенли и др., 1973).
В опытах с импульсной меткой и последующим продолжительным вытеснением ее немеченым субстратом показано, что все прочие основные кодируемые вирусом белки совершенно стабильны и не являются продуктами посттрансляционного расщепления каких-либо высокомолекулярных предшественников. (Тейлор и др., 1969).
Репликация РНК ортомиксовирусов
Свыше 80% РНК, синтезирующейся в присутствии актиномицина D, после третьего часа инфекции представляет собой вРНК. (Дьюсберг и Робинсон, 1967; Нейак и Балуда, 1968; Шолтиссек и Ротт, 1970). Очевидно, посредством какого-то регуляторного механизма должны создаваться условия, благоприятствующие предпочтительному синтезу вРНК, а не кРНК, несмотря на то что последняя необходима как для трансляции, так и для репликации.
Понс (1970) предположил, что индивидуальные молекулы РНК могут оставаться во время репликации связанными, возможно располагаясь одна за другой на остове, состоящем из молекул белка NP, однако это предположение еще не доказано. Нейак и Балуда (1968) описали структуру, которая, вероятно, представляет собой репликативный предшественник: это быстро метящаяся 14S-PHK, частично устойчивая к РНКазе и осаждаемая 2М NaCl. Репликаза до сих пор не индентифицирована; неизвестно также, где происходит репликация (равно как и транскрипция) — в ядре или в цитоплазме.
- Читать далее "Сборка ортомиксовирусов. Выход ортомиксовирусов из клетки"
Оглавление темы "Характеристика тогавирусов и ортомиксовирусов":1. Метаболизм клеток пораженных пикорнавирусами. Синтез белков пораженными пикорнавирусами клетками
2. Подавление синтеза клеточных РНК пикорнавирусами. Подавление синтеза ДНК пикорнавирусами
3. Дефектные частицы пикорнавирусов. Цикл размножения вируса ящура и тогавирусов
4. Трансляция тогавирусов. Этапы трансляции тогавирусов
5. Репликация РНК тогавирусов. Транскрипция РНК тогавирусов
6. Сборка и выход тогавирусов из клетки. Метаболизм пораженной тогавирусами клетки и флавивирусы
7. Размножение вируса краснухи в клеточных культурах. Цикл размножения ортомиксовирусов
8. Начало ортомиксовирусной инфекции. Транскрипция ортомиксовирусов
9. Трансляция ортомиксовирусов. Репликация РНК ортомиксовирусов
10. Сборка ортомиксовирусов. Выход ортомиксовирусов из клетки