Сборка ортомиксовирусов. Выход ортомиксовирусов из клетки
Сборка вирионов гриппа происходит одновременно с их высвобождением из клетки посредством почкования (ДакНгуен и др., 1966). Этот процесс был детально изучен Компансом и Диммоком (1969). Участки плазматической мембраны, содержащие гемагглютинин и нейраминидазу, образуют видимые выросты примерно в то же самое время, когда на внутренней стороне мембраны формируется новый слой белка (предположительно полипептида М). К этим морфологически измененным зонам мембран присоединяется рибонуклеопротеид (РНП) и происходит почкование сферических или нитевидных вирионов. Данный процесс будет более подробно обсуждаться в разделе, посвященном парамиксовирусам, однако два вопроса имеет смысл рассмотреть именно здесь: состояние, в котором находится РНП, и роль нейраминидазы.
Выделенные из зараженных клеток РНП — это короткие образования, длина которых соответствует длине входящих в их состав молекул РНК (Дьюсберг, 1969; Кингсбери и Уэбстер, 1969; Понс и др., 1969). Структурно они сходны с РНП, выделенными непосредственно из вирионов, т.е. представляют собой двойную спираль с одноцепочечной петлей на одном из концов.
Компанс и др. (1970а) рассчитали, что вероятность точной комбинации 5 (а сейчас предполагают, что 7 или более) фрагментов РНП, случайно отбираемых из внутриклеточного пула для включения в почкующийся вирион, неправдоподобно мала, если не предположить, что вирион в среднем содержит избыток таких фрагментов.
Вместе с тем не исключено, что в нативном состоянии в зараженных клетках и вирионах РНП представляет собой одну непрерывную нить, включающую полный набор молекул РНК, и что описанные выше фрагменты РНП — результат разрыва этой структуры в слабых точках между индивидуальными молекулами РНК (Понс, 1970).
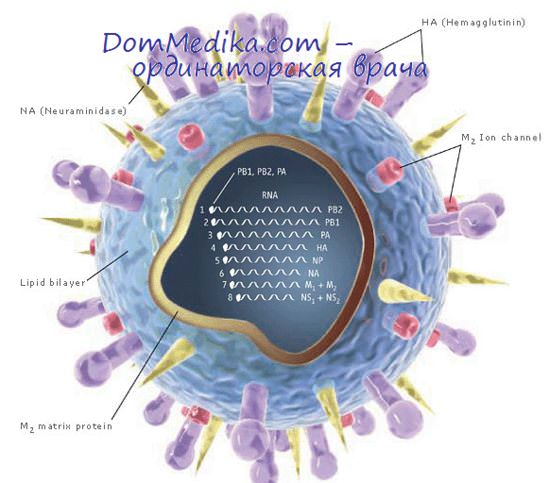
Нейраминидаза попадает в определенные районы плазматической мембраны приблизительно в то же время, что и гемагглютинин, причем эти районы часто расположены около одного из полюсов распластанных в монослое клеток (Мейно и Килборн, 1970). Без сомнения благодаря этому ферменту нейраминовая кислота в гликопротеидах и гликолипидах этих районов мембран отсутствует (Кленк и др., 1970).
Все же кажется маловероятным, что нейраминидаза играет существенную роль при включении гликопротеидов в мембраны, образовании пепломеров или самом почковании, так как многие почкующиеся вирусы вообще лишены этого фермента. Однако нейраминидаза, возможно, принимает участие в облегчении элюирования почкующихся вирионов с наружной поверхности клетки (Компанс и др., 1970а), так как антитела против нейраминидазы (но не моновалентные фрагменты Fab, полученные в результате обработки пепсином) подавляют высвобождение вируса (Бехт и др., 1971).
Антигемагглютинин действия не оказывает, но конканавалин А, который, по-видимому, сильнее связывается с нейраминидазой, чем с гемагглютинином, блокирует образование вируса (Ротт и др., 1972).
Количества образующихся вирусных структурных белков не только не эквимолярны, но и далеки от тех соотношений, в которых эти белки представлены в вирионе (Уайт и др., 1970). Вероятно, выражение генов белков NA и Р должно подавляться с помощью некоего регуляторного механизма, поскольку эти белки едва определимы в зараженных клетках, тогда как полипептиды NP и NS образуются в очень больших количествах.
Синтез двух последних полипептидов выявляется уже на втором часу после заражения и затем увеличивается, выходя на плато между 5-м и 8-м часами. Скехел (1972) обнаружил, что относительные скорости синтеза вирусных белков меняются во времени; на ранних сроках цикла в клетках много полипептида NS и мало полипептида М, на поздних соотношения этих компонентов обратные.
- Вернуться в оглавление раздела "Вирусология"
Оглавление темы "Характеристика тогавирусов и ортомиксовирусов":1. Метаболизм клеток пораженных пикорнавирусами. Синтез белков пораженными пикорнавирусами клетками
2. Подавление синтеза клеточных РНК пикорнавирусами. Подавление синтеза ДНК пикорнавирусами
3. Дефектные частицы пикорнавирусов. Цикл размножения вируса ящура и тогавирусов
4. Трансляция тогавирусов. Этапы трансляции тогавирусов
5. Репликация РНК тогавирусов. Транскрипция РНК тогавирусов
6. Сборка и выход тогавирусов из клетки. Метаболизм пораженной тогавирусами клетки и флавивирусы
7. Размножение вируса краснухи в клеточных культурах. Цикл размножения ортомиксовирусов
8. Начало ортомиксовирусной инфекции. Транскрипция ортомиксовирусов
9. Трансляция ортомиксовирусов. Репликация РНК ортомиксовирусов
10. Сборка ортомиксовирусов. Выход ортомиксовирусов из клетки