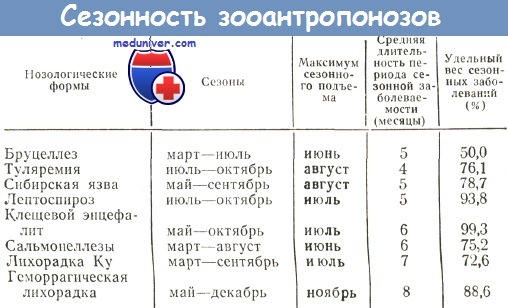
Эпизоотологическая эффективность переносчика инфекции и его характеристика
Эпизоотологическая эффективность переносчика характеризуется как функция следующих факторов:
1. Количество видов, участвующих в переносе возбудителя в данной местности в данное время.
2. Численность переносчиков каждого вида в данной местности в данное время.
3. Интенсивность контакта с соответствующим хозяином или, если в естественный цикл вовлекается более одного вида позвоночных, со всеми возможными хозяевами.
4. Алиментарная активность.
5. Способность к восприятию возбудителя и последующей передаче его позвоночному хозяину.
Влияние географической среды, а также деятельности человека, сказывается на эпизоотологической эффективности переносчиков возбудителей через один из указанных факторов.
Список видов и форм блох, из которых выделяли культуры возбудителя чумы, превышает 99 названий, не считая блох, паразитирующих на крысах и домовых мышах (Фенюк). В передаче возбудителя сибирской язвы могут принимать участие представители 17 видов клещей, 9 видов жалящих и лижущих мух, 3 вида комаров. Однако основная роль в трансмиссивной передаче возбудителей сибирской язвы в определенных ландшафтах принадлежит слепням или комарам (Черкасский и Лаврова).
На территории России к настоящему времени установлена естественная зараженность возбудителем туляремии 20 видов иксодовых и 10 видов гамазовых клещей, 17 видов блох, 11 видов комаров, 6 видов слепней, а также мошек, клопов, вшей (Олсуфьев и Дунаева). Несмотря на это, основными переносчиками возбудителя туляремии могут быть признаны лишь иксодовые клещи и кровососущие двукрылые, причем первые являются не только переносчиками, но и длительными хранителями возбудителя инфекции в межэпизоотические периоды.
Аналогичное явление характерно и для многих других трансмиссивных зооантропонозов. В связи с этим возникла необходимость дифференцировать переносчиков на основных и второстепенных.
Численность популяции членистоногих имеет решающее значение и зависит от нескольких факторов, влияющих на воспроизведение и продолжительность их жизни. Наиболее важны благоприятные климатические условия, при которых срок жизни членистоногих удлиняется. Для комаров и других двукрылых насекомых наличие благоприятных мест выплода, а для клещей и других эктопаразитов — многочисленность подходящих хозяев в значительной мере определяют численность популяции членистоногих.
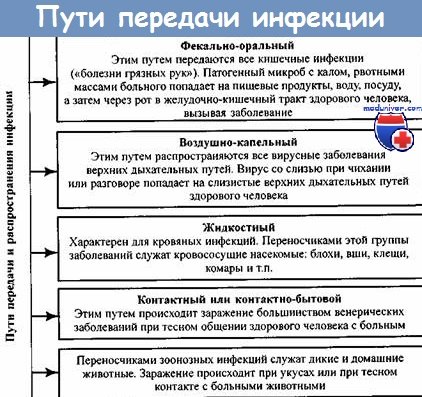
По характеру соотношений с передаваемыми возбудителями и по механизму самой их передачи переносчиков подразделяют на механических и специфических.
Механические переносчики осуществляют заражение контактным путем — через инокуляцию возбудителя зараженными ротовыми органами или через контаминацию реципиента инфицированными испражнениями (перенос возбудителя сибирской язвы клещами и др.).
Возбудитель может также проникнуть через слизистую конъюнктивы глаза или через дыхательные пути. Так, вирус энцефалита Западного Нила может выделяться с экскретами клеща Ornithodorus spp. и поэтому возможна передача путем контаминации.
Механический способ переноса — первая стадия приспособления возбудителя к переносчику. Более совершенной в биологическом аспекте и более эффективной в эпизоотологическом отношении следует считать специфическую передачу возбудителей кровососущими членистоногими.
В процессе переноса возбудителя специфическими переносчиками возбудитель в организме членистоногого продолжает паразитический образ жизни, проходя часть своего биологического цикла (филяриозы, лейшманиозы) или многократно размножаясь (клещевые энцефалиты, клещевые спирохетозы).
Среди специфических переносчиков могут наблюдаться:
а) пропагативная форма (непосредственное размножение возбудителя в переносчике);
б) циклометаморфозная форма (возбудитель проходит в переносчике часть своего жизненного цикла без размножения);
в) циклопропагативная форма (возбудитель, проходя в переносчике жизненный цикл, размножается).
Передача возбудителей при специфическом переносе осуществима не ранее, чем возбудители, попавшие в организм переносчика, пройдут сложный процесс адаптации, развития или размножения до количества, достаточного для последующего заражения позвоночного животного, и переместятся в организме переносчика в такое место, откуда возможен выход наружу (слюнные железы, эпителий кишечника и т. п.).
Для заражения каждого вида членистоногого требуется определенное количество данного возбудителя в крови позвоночного хозяина. Количественно его выражают как титр возбудителя в крови позвоночного хозяина, необходимый и достаточный для заражения 50% питавшихся этой кровью членистоногих.
Говоря об интенсивности контактов переносчиков с биологическими хозяевами возбудителя, следует рассмотреть два аспекта этого вопроса: привязанность представителей данного вида переносчиков к определенным группам животных или птиц и вид паразитизма переносчиков по отношению к хозяину возбудителя. Так, род Culex тесно связан с различными дикими и домашними птицами, хотя некоторые его виды способны нападать и на млекопитающих. Это обусловливает передачу комарами этого рода «птичьих» вирусов японского энцефалита, лошадиного энцефалита, энцефалита Западного Нила и др.
Комары Aedes охотнее нападают на млекопитающих, в том числе и на мелких грызунов, что определяет их участие в передаче возбудителя туляремии среди водяных полевок и крыс. Другие представители этого рода, обитающие в кронах деревьев тропических лесов, распространяют вирусы желтой лихорадки среди обезьян. Род Anopheles связан в основном с крупными млекопитающими.
В то же время, например, слепни являются паразитами, не приуроченными к каким-либо определенным видам хозяев, хотя и нападают преимущественно на крупных млекопитающих.
Членистоногие, привязанные к определенному виду хозяев, могут быть более эффективными в эпизоотологическом отношении, чем многохозяинные.
Характер общения переносчика с хозяином возбудителя оказывается различным у разных видов кровососущих членистоногих. Так, комары и слепни связаны с хозяевами лишь факультативно и места их обитания обычно не соотносятся с поселениями последних. Клещи (гнездово-норовые паразиты) контактируют с хозяином только в момент кровососания, а весь цикл развития (спаривание, переваривание крови, отклад яиц, линька) протекает в гнезде или норе (например, «блохи гнезда» по Иоффу, большинство гамазид).
Пастбищные подстерегающие паразиты также контактируют с хозяином лишь в период кровососания, однако заселяют не только убежище хозяина, но и другие посещаемые им места (иксодовые клещи).
Не менее важным условием эффективности членистоногих как переносчиков возбудителей инфекций являются особенности их взаимоотношений с возбудителем. Так, феномен «чумного блока» у блох, являющийся обязательным и строго специфичным фактором в механизме передачи возбудителей чумы, далеко не всегда и не в одинаковой мере свойствен различным родам блох. Экспериментально показано, что сроки блокообразования у блох рода Xenopsylla колеблются от 6 до 55 дней, тогда как у рода Neopsylla они достигают 110 дней, возникновение блока у Oropsylla silantiewi и С. lebedevi наблюдали между 4 и 51-м днями, а у Rhadinopsylla liventricosa — лишь на 16—93-й день после заражающего кормления (Флегонтова, Шварц и др.). В то же время у блох мышей — L. segnis, а также у блох рода Ctenophalmus блокообразование практически не происходит (Флегонтова, Kartman et al.).
Клещи Ornithodorus, как известно, являются специфическими переносчиками возбудителя клещевого опирохетоза. Кроме того, они легко воспринимают возбудителей бруцеллеза, чумы, туляремии, лихорадки Ку, однако случаи успешной передачи ими возбудителей этих инфекций через укус биопробным животным чрезвычайно редки.
В то же время если для возбудителей клещевого реккуренса организм клещей Ornithodorus papillipes и ряда других видов является специфической средой обитания, то в О. lahorensis, Argas persicus и других этот микроорганизм погибает, хотя все эти эктопаразиты обитают в норах одних и тех же животных, служащих для них источником питания.
Фактором, сдерживающим распространение возбудителей инфекций членистоногими переносчиками, является нередко наблюдаемая частичная, а подчас и полная гибель возбудителей в организме переносчика.
Гибель палочки чумы в организме блохи, возможно, связана с воздействием бактерицидных веществ кишечника блох, фагоцитозом, воздействием бактериофага, а также питанием зараженных блох кровью иммунных к чуме животных (Бибикова). Частичную гибель возбудителей клещевого риккетсиоза в клещах Н. asiaticum и О. lahorensis также объясняют воздействием бактерицидных веществ, содержащихся в кишечнике клещей (Гроховская, Сидоров).
Алиментарная активность кровососущих членистоногих, являющаяся видовым свойством, но регулируемая условиями существования, во многом определяет эпизоотологическую эффективность переносчиков, поскольку высокая частота кровососания не только увеличивает вероятность заражения большего количества животных, но и оказывает стимулирующее влияние на размножение микробов в переносчике (например, возбудителя туляремии в клещах, возбудителя чумы в блохах и т. п.).
Климатотеографические условия существенно влияют на эпизоотологическую эффективность переносчиков возбудителей инфекций, определяя их ареал и численность, стимулируя или подавляя их жизнеспособность, регулируя биологические взаимоотношения переносчика с возбудителем. Так, у иксодовых клещей продолжительность развития личинок в нимф составляет 16—18 дней при температуре 15—17°С и 10 при температуре 27—28°С. Активность иксодовых клещей, выраженная в движении, обонянии, способности цепляться проявляется уже при температуре 0,3—1°С, тогда как при температуре свыше 20°С клещи становятся менее активными и переходят с растительности в подстилку (Пионтковская, Шмаева). Полеты слепней и нападения их на животных начинаются при температуре воздуха не ниже 15—16°С, становятся интенсивными при 19—20°С и выше, а температура выше 31—32°С действует на слепней угнетающе.
Термофильность сочетается у слепней с большой чувствительностью к недостатку влаги: если подержать слепней несколько часов без увлажнения при температуре 25—30°С, они погибнут (Олсуфьев).
Вирус японского энцефалита размножается в комарах-переносчиках при температуре не ниже 20—25°С, причем температура 25—35°С ускоряет размножение. Комары-переносчики вируса желтой лихорадки приобретают способность передавать возбудителя при температуре воздуха 21°С через 18 дней, при 36—37°С — через 4—5 дней, а при температуре ниже 18°С размножение вируса в организме комара прекращается (Петрищева). Сроки образования блохами чумного блока и, следовательно, способность передавать возбудителей также зависят от температуры окружающей среды.
Так, при 8°С блохи X. g. minax блокируются в среднем через 10,6 дня, при 22°С — через 8,2 дня, при 28°С — через 6,1 дня, в блохах X. cheopis при 8°С блок образуется в среднем за 34,4 дня, при 21°С — за 12,6 и при 30°С — за 11,9 дня (Бибикова).
На эпизоотологическую эффективность переносчиков возбудителей инфекций может влиять как обычная хозяйственная деятельность человека, так и проведение специальных мероприятий, причем влияние это прежде всего сказывается на численности кровососущих членистоногих.
Примером может служить освоение тайги, в результате которого дикие копытные животные уходят или истребляются, что может вести к гибели клещей-переносчиков возбудителя клещевого энцефалита. Гидромелиоративные мероприятия, направленные на осушение водоемов или заболоченных территорий, лишают комаров мест выплода, что ведет к резкому уменьшению их численности и т. п.
В то же время хозяйственная деятельность человека может невольно вести и к увеличению численности переносчиков, что, например, наблюдается в отношении клещей на участках леса, служащих для выпаса домашнего скота, на который и переходят кормиться клещи— переносчики возбудителей болезней.
При разных зооантропонозах возможен разнообразный характер течения эпизоотического процесса — как без межэпизоотического периода, так и с различными вариантами протекания его с сохранением возбудителя в переносчиках или на неживых объектах внешней среды.
Эпизоотический процесс при лептоспирозе может неопределенно долго поддерживаться и среди сельскохозяйственных животных, чему способствует обильное и длительное выделение лептоспир с мочой животных, достаточно длительный срок выживания возбудителя, особенно во влажной среде, многообразие путей проникновения лептоспир в организм животных, а также крупногрупповое содержание животных, обеспечивающее тесноту их общения.
В эпизоотологии бруцеллеза при бессимптомном и латентном течении инфекции животное служит бруцелловыделителем и может играть роль источника инфекции, остающегося вне подозрения. Аборты усугубляют опасность бруцеллезных животных, принимая иногда массовый характер и охватывая до 70% маточного поголовья. В период абортов резко повышается интенсивность выделения бруцелл во внешнюю среду с патологическими истечениями из родовых путей, которые могут длиться до 15 месяцев. Клинически выздоровевшие животные могут оставаться бактерионосителями в течение 16—20 месяцев, иногда 3—4 года, а отдельные животные — в течение 5 и более лет.
Сходная картина может наблюдаться при листериозе. Селькохозяйственные животные (крупный и мелкий рогатый скот, свиньи, лошади) выделяют возбудителей листериоза во внешнюю среду с истечениями из носовой полости, из половых органов (при абортах), с абортированным плодом, с калом, мочой, молоком. Среди животных нередко бессимптомное листерионосительство. Доказана стойкость листерий во внешней среде и даже способность их размножаться в воде и почве. В силу этого листериоз в животноводческом хозяйстве может существовать в течение трех и более лет, без четко выраженного межэпизоотического периода.
При лимфацитарном хориоменингите вирус хранится в обыкновенных (домовых) мышах и других мелких грызунах с постоянной вирусемией, не вызывающей, однако, их гибели. Хотя передача возбудителей среди животных осуществляется лишь посредством переносчиков (гамазовых клещей), при этой инфекции межэпизоотический период вообще отсутствует.
Можно считать доказанной способность сохранения возбудителя в зимний межэпизоотический период в переносчиках и при некоторых других трансмиссивных зооантропонозах. Доказана способность комаров С. fatigans сохранять вирус энцефалита Сан-Луи в течение 116 дней, в том числе и в зимний период. На основании фактов выделения вируса западного лошадиного энцефалита из зимующих комаров С. tarsalis признано, что комары могут заразиться перед уходом на зимовку и сохранять вирус в течение диапаузы. Имеются указания о способности зимовать в комарах и вируса японского энцефалита.
Наряду с этим межэпизоотический период может длиться в течение нескольких, иногда многих лет.
Способность возбудителей не только длительно сохраняться, но и при определенных условиях размножаться на объектах внешней среды, как это имеет место при сибирской язве, столбняке, листериозе и некоторых других, обеспечивает их сохранение в течение неопределенно долгого межэпизоотического периода.
В большинстве случаев не представляет труда объяснить переживание в течение длительного межэпизоотического периода возбудителей, передающихся клещами. Клещи могут сохранять в себе, например, туляремийных бактерий пожизненно.
Наличие периодов длительного затишья (10 и более лет), за которыми следовали внезапные вспышки чумы у грызунов или людей, неоднократно подтверждалось в некоторых природных очагах. Были выдвинуты различные объяснения повторного проявления чумы на одной и той же территории после длительных периодов затишья.
Так, по мнению Н. И. Калабухова, без ежегодно протекающих эпизоотии невозможно поддержание эпизоотии чумы. В межэпизоотический период условия для циркуляции микроба отсутствуют, а возникновение новых эпизоотии — результат заноса чумы из смежных эпизоотических районов.
Показано, что чумной микроб способен длительно выживать в почве нор грызунов. Микроба выделяли из почвы даже тех нор, в которых грызуны погибли по крайней мере за 11 месяцев до этого. Здоровые грызуны, занимающие такие норы, могут заразиться путем контакта с зараженными слоями почвы. Как уже говорилось, на основе подобных фактов М. Baltazard сформулировал теорию о теллурической («рытьевой») чуме, согласно которой микроб сохраняется в межэпизоотический период в почве нор в «непаразитарной стадии».
В последнее время все больше сторонников приобретает теория микроочагового сохранения чумы в неэпизоотические годы в виде цепочки спорадических случаев. Чрезвычайно малая в этих условиях вероятность обнаружения возбудителя чумы при обычных нормах эпизоотологического обследования позволяет говорить о межэпизоотическом периоде.
- Читать "Современная классификация зооантропонозов"
Оглавление темы "Эпидемиология зооантропонозов":- Воздушный механизм передачи возбудителя инфекции при зооантропонозах
- Трансмиссивный механизм передачи возбудителя инфекции при зооантропонозах
- Контактный механизм передачи возбудителя инфекции при зооантропонозах
- Восприимчивость человека к зоонозам - зооантропонозам
- Эпидемиология зооантропонозов и пути их распространения
- Типы заболеваемости зооантропонозами - бытовые, профессиональные
- Эпидемиология зооантропонозов в России и СССР
- Эпизотии зооантропонозов
- Эпизоотологическая эффективность переносчика инфекции и его характеристика
- Современная классификация зооантропонозов