Реакция Т-лимфоцитов на ВИЧ-инфекцию
При большинстве вирусных инфекций Т-клеточный иммунный ответ играет решающую роль в подавлении или элиминации инфекционного агента. В случае ВИЧ противовирусная активность включает в себя как механизмы неспецифической резистентности к инфекциям («врожденный иммунитет»), так и иммунный ответ (т.е. реакции собственно иммунной системы).
В данной статье рассматриваются реакции Т-лимфоцитов, направленные против ВИЧ, которые осуществляются через распознавание вирусных белков либо в составе вирусных частиц, либо в составе инфицированных ВИЧ клеток. Поскольку ВИЧ-инфекция поражает и саму иммунную систему, некоторые данные, представленные в Главе 8, также имеют прямое отношение к обсуждаемой теме. Кроме того, здесь описана нецитотоксическая активность CD8+-клеток в отношении ВИЧ, которая относится к реакциям врожденного иммунитета.
Другие клеточные реакции системы врожденного иммунитета против ВИЧ со стороны NK-клеток, NK-T-клеток и Тγδ-клеток описаны в отдельных статьях на сайте. Важно отметить, что эффективность врожденных и приобретенных клеточных иммунных реакций зависит также от эффективной работы антигенпредставляю-щих клеток (АПК), в особенности макрофагов и дендритных клеток.
Существует две основные категории CD4+- и CD8+-Т-лимфоцитов: не контактировавшие с антигеном (неиммунные) и контактировавшие с антигеном (иммунные) клетки памяти. Т-клетки памяти представляют собой фенотипически гетерогенную популяцию, которую можно подразделить на два основных типа.
(i) Центральные Т-клетки памяти (Тцп), циркулирующие во вторичных лимфоидных тканях через кровь и лимфатическую систему. Эти клетки экспрессируют CCR7 и CD62L, что позволяет им проникать в лимфоидную ткань и рециркулировать.
(ii) Эффекторные Т-клетки памяти (Тэп), мигрирующие из вторичных лимфоидных тканей в экстралимфоидные ткани, в частности в кишечник (базальную пластинку и эпителий).
Эффекторные Т-клетки памяти не экспрессируют CCR7 и CD62L, могут активно реагировать на антиген (в том числе на ВИЧ) и превращаться в действующие эффекторные клетки; эти клетки после исчезновения антигена существуют в течение примерно 1 недели или превращаются в центральные Т-клетки памяти. Некоторые исследователи используют термин «покоящиеся» по отношению к центральным клеткам памяти, чтобы подчеркнуть отличие их размеров (небольшие) по сравнению с активированными клетками памяти и отсутствие поверхностных клеточных маркеров (например, HLA-DR, CD69).
Покоящиеся клетки памяти, таким образом, представляют собой Т-лимфоциты, которые встречались с антигеном, как и Тэп-клетки, но при уменьшении количества антигена могут возвращаться в состояние покоя. Они способны вновь превращаться в эффекторные клетки памяти после повторного контакта с антигеном. Таким образом, покоящиеся клетки памяти обладают характеристиками Тцп-клеток.
Тцп-клетки после повторного контакта с антигеном быстро начинают вырабатывать цитокины. У них снижается экспрессия CCR7, они начинают пролиферировать и перемещаются в периферические органы, превращаясь в Тэп-клетки. Последние остаются в месте проникновения инфекции и защищают организм. При контакте с антигеном Тэп-клетки активируются и превращаются в эффекторные клетки, а затем в короткоживущие терминальные эффекторные клетки.
Некоторые из них могут обратно переходить в состояние Тцп (покоящееся памяти). Очевидно, что Тцп- и Тэп-клетки (особенно CD8+-клетки) могут менять свой функциональный фенотип в обе стороны. Тэп-клетки обладают ограниченной способностью к размножению, тогда как наивные и Тцп-клетки живут дольше и могут проходить через большое количество циклов деления. Так, некоторые исследования показали персистенцию вакцинальных вирусспецифических CD4+-Tцп клеток в течение 70 лет после вакцинации.
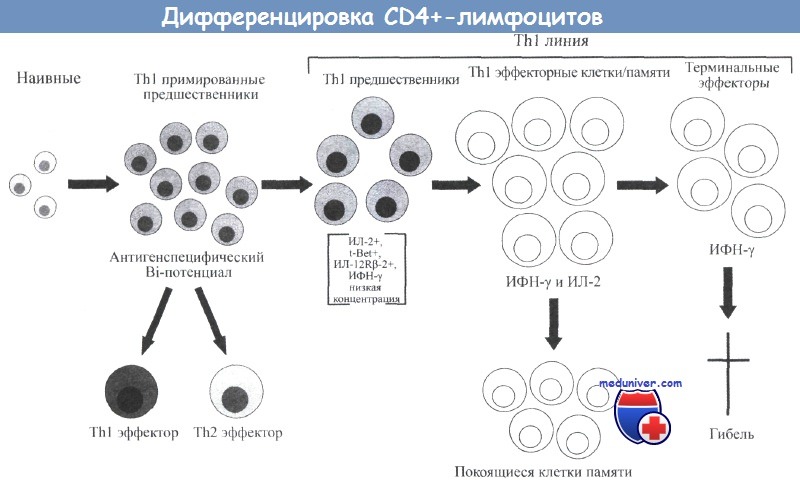
Общее развитие различных субпопуляций Т-клеток также отражается в формировании субпопуляций CD8+-Т-клеток.
Наивные клетки после контакта с антигеном превращаются в эффекторные клетки, которые в случае CD4+-клеток имеют Тh1 или Тh2 фенотип.
Эффекторные клетки могут превращаться в эффекторные клетки (памяти) или покоящиеся (центральные) клетки памяти в присутствии низких концентраций антигена.
При повторном контакте с антигеном эффекторные Т-клетки памяти быстро реагируют и превращаются в терминальные эффекторные клетки.
Эти клетки являются короткоживущими, обладают высокой специфичностью по отношению к антигену и проявляют цитотоксическую активность, которая особенно характерна для CD8+-клеток.
CD8+- и CD4+-Т-лимфоциты, проявляющие активность цитотоксических Т-лимфоцитов (ЦТЛ), обычно реагируют на презентацию эпитопов АПК в комплексе с молекулами МНС I или II класса соответственно. Антиген распознается с помощью Т-клеточного рецептора (TCR), представляющего собой гетеродимерную клеточную поверхностную молекулу, которая взаимодействует с чужеродным антигеном, презентируемым в виде коротких пептидов вместе с молекулами МНС I или II класса на поверхности клетки.
TCR состоит из а-, b-, у- или 8-цепей, хотя подавляющее большинство циркулирующих Т-лимфоцитов экспрессирует ар-цепи (описание Тγδ-клеток см. в отдельнной статье на сайте). Распознавание вирусных патогенов с молекулами главного комплекса гистосовместимости I класса, по-видимому, является более точным процессом не только за счет ограниченной длины пептидов (от 8 до 10 аминокислотных остатков), но и в связи с тем, что концевые участки пептидов (карбоксиконцевой и аминоконцевой) должны соответствовать определенным выемкам на противоположных сторонах углублений в локусе МНС I класса.
Пептиды МНС II класса могут быть длиннее (от 12 до 24 остатков) и, таким образом, обладают большей гетерогенностью. В связи с этим разнообразие антигенных детерминант, презентируемых CD4+-клеткам с молекулами МНС II класса, может быть больше по сравнению с детерминантами, презентируемыми CD8+-клеткам с молекулами I класса.
Генетическая основа человеческих лейкоцитарных антигенов (HLA) (т.е. МНС) и факторы среды могут влиять на репертуар созревающих Т-клеток, разнообразие которых может быть выгодно для организма хозяина. Нарушения в репертуаре Т-клеток (что выражается в использовании тех или иных сегментов генов при перестройке ДНК вариабельных областей антигенсвязывающих субъединиц TCR) также могут наблюдаться при ВИЧ-инфекции в случае:
(i) экспрессии других вирусных антигенов, которые вызывают селективную экспансию и гибель клеток,
(ii) молекулярной мимикрии фрагментов gp120, сходных с HLA,
(iii) встраивания молекул HLA в оболочку вируса и (iv) суперантигенных эффектов ВИЧ.
Общим результатом является дисбаланс в разнообразии реакций CD8+- или CD4+-клеток, который может приводить к неблагоприятным последствиям.
При ограничении репертуаров TCR на CD4+-и CD8+-лимфоцитах иммунный ответ может быть неполноценным, особенно в ответ на попадание в организм новых антигенов. Таким образом, степень разнообразия репертуара Т-клеточных реакций во время первичной инфекции может являться прогностическим признаком того или иного клинического исхода заболевания. В частности, при исследовании ВИЧ-инфицированных людей была показана возможность клональной или олигоклональной экспансии CD4+- и CD8+-клеток.
Поскольку экспансии разнах субпопуляций лимфоцитов не обязательно происходят скоординированно, могут возникать расхождения или «разрывы» в репертуарах разных субпопуляций Т-клеток, которые способны увеличиваться по мере прогрессирования заболевания. Тем не менее и в этих случаях возможна некая пластичность в развитии иммунного ответа.
Нарушение репертуара реагирующих лимфоцитов может приводить к появлению как положительных, так и отрицательных последствий, если клетки проявляют активность ЦТЛ или осуществляют другие реакции, направленные против ВИЧ. Отсутствие или уменьшение определенных специфичностей Т-клеток может уменьшать сопротивляемость инфекциям и способствовать развитию злокачественных опухолей.
Также необходимо отметить, что, несмотря на изменение репертуара Т-клеток в периферической крови, их распределение в лимфоидной ткани (например, в миндалинах) может оставаться неизменным. Это, возможно, является причиной того, что уменьшение количества CD4+-Т-клеток в периферической крови может не сопровождаться аналогичным уменьшением таких клеток в периферических лимфоидных тканях, за исключением, вероятно, выраженного уменьшения количества Т-лимфоцитов в слизистой оболочке желудочно-кишечного тракта.
Ранние исследования показали возможность существования физиологической регуляции Т-лимфоцитов в организме хозяина. Потеря CD4+-лимфоцитов, по-видимому, компенсируется за счет увеличения количества CD8+-лимфоцитов. CD8+-клеточный лимфоцитоз может влиять на регенерацию CD4+-клеток. Если такой гомеостатический механизм существует, можно разработать методы увеличения количества CD4+-клеток у ВИЧ-инфицированных лиц за счет изменения количества CD8+-клеток.
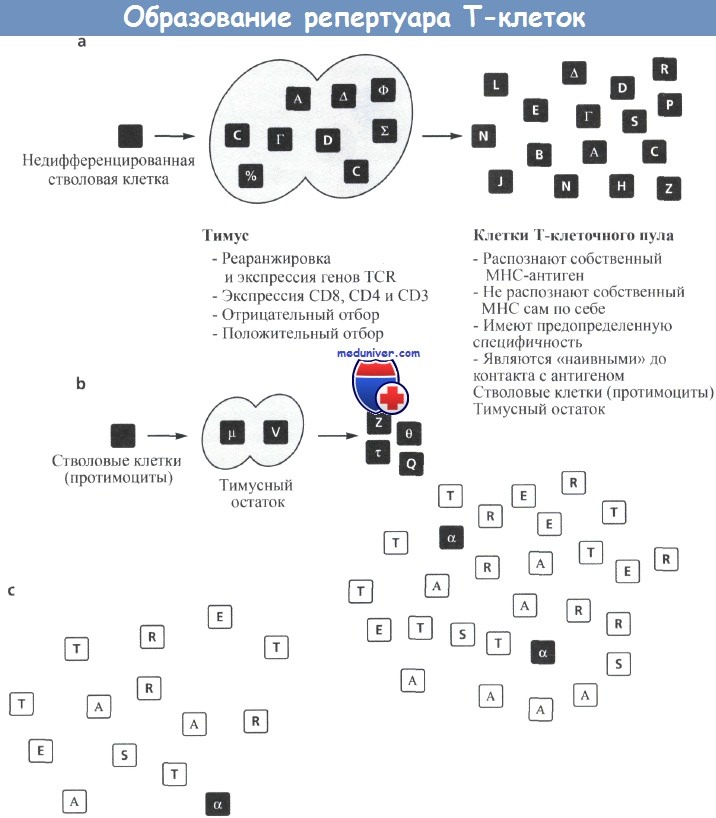
(а) Недифференцированные лимфоидные стволовые клетки (протимоциты), созревая в тимусе, начинают экспрессировать молекулы CD4 и CD8.
Гены их Т-клеточных рецепторов (TCR) подвергаются перестройке и начинают экспрессироваться, а образовавшиеся в результате тимоциты подвергаются отрицательной или положительной селекции. Клоны Т-клеток, выходящие в циркуляцию, обладают различными TCR, что указывает на их потенциал для осуществления поликлональных реакций.
(b) При разрушении Т-клеток под действием ВИЧ может сохраняться ограниченный спектр Т-клеток, при регенерации которого на фоне различных терапевтических воздействий может не происходить восстановления первоначального репертуара, присутствовавшего в организме хозяина,
(с) В конечной стадии спектр Т-клеток может быть настолько ограничен, что не дает им возможности быстро реагировать на определенный антиген. В некотором смысле, как показано на рисунке, репертуар Т-клеток не сможет реагировать так, как это требуется для распознавания точной последовательности ZEBRA.
Тем не менее они могут отвечать с помощью других механизмов (хотя и более медленно), распознавая антиген как «лошадь с черными и белыми полосками».
- Читать "Методы оценки реакции Т-лимфоцитов на ВИЧ-инфекцию"
Оглавление темы "Иммунный ответ на ВИЧ-инфекцию":- Комплементсвязывающие антитела к ВИЧ
- Аутоиммунные реакции и болезни при ВИЧ
- Пролиферация В-клеток при ВИЧ
- Молекулярная мимикрия при ВИЧ
- Антиидиопатические антитела при ВИЧ
- Резюме по гуморальному иммунитету у больных ВИЧ
- Реакция Т-лимфоцитов на ВИЧ-инфекцию
- Методы оценки реакции Т-лимфоцитов на ВИЧ-инфекцию
- Классификация CD4 Т-лимфоцитов
- Реакции CD4 Т-лимфоцитов