Трансляция и репликация РНК парамиксовирусов. Сборка и выход парамиксовирусов из клетки
Пять или шесть полипептидов, обнаруженных в вирионах парамиксовирусов, выявляются также на электрофореграммах препаратов из зараженных клеток. Два из них содержат углеводы, половина которых представлена глюкозамином, а остальная часть — галактозой, маннозой и фукозой (Кленк и др., 1970а; Чоппин и др., 1971). Неструктурные белки, индуцированные вирусом, пока не описаны.
С помощью методов иммунофлуоресценции (Реда, 1964) и электронной микроскопии (Чоппин и др., 1966) в цитоплазме обнаружены большие агрегаты РНП. При заражении клеток некоторыми парамиксовирусами, особенно вирусами подгруппы кори — чумы рогатого скота — чумы плотоядных, этими же методами или даже с помощью общепринятых методик окрашивания выявлены ядерные включения; однако накапливающийся в ядрах белок, вероятно, не играет существенной роли в размножении вирусов (Льянес-Родас и Лиу, 1965).
Репликативный предшественник (РП) до сих пор достоверно не обнаружен. Портнер и Кингсбери (1972) описали медленно метящиеся 24S-PHK, практически полностью устойчивые к РНКазе и при плавлении дающие одноцепочечные РНК 50S, которые гибридизуются с вРНК несколько эффективнее, чем молекулы последней гибридизуются друг с другом. Не исключено, что эта РНК — по мнению авторов РП— на самом деле является репликативной формой. Фермент, катализирующий репликацию РНК, пока не выделен.
Портнер и Кингсбери (1971) описали дефектные (неинфекционные) вирионы Сендай, которые содержали только короткие молекулы РНК и препятствовали размножению вируса Сендай (но не других парамиксовирусов), подавляя скорее репликацию, чем транскрипцию вирусной РНК- Механизм этого явления, возможно, сходен с аутоинтерференцией, характерной для Т-частиц вируса везикулярного стоматита, которые подробнее обсуждаются в статье, посвященном рабдовирусам.
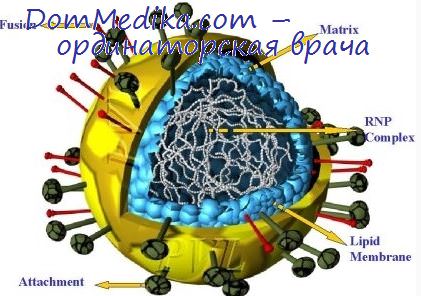
Сборка и выход парамиксовирусов из клетки
Сборка парамиксовирусов всесторонне изучена в лаборатории Чоппина (см. обзоры: Чоппин и др., 1971; Компанс и Чоппин, 1971). Первым этапом сборки является включение вирусных гликопротеидов в плазматическую мембрану; к участкам мембраны, в которые включаются гликопротеиды, способны прилипать эритроциты или конъюгированные с ферритином антитела. Затем в зонах, где происходит почкование, проявляются морфологические изменения трех типов: 1) гликопротеиды приобретают свою характерную шиловидную структуру; 2) на внутренней стороне плазматической мембраны под шипами появляется электроноплотный слой (вероятно, состоящий из белка М); 3) нуклеокапсиды располагаются вдоль утолщенных зон мембраны. Почкование происходит только в тех участках, в которых обнаруживаются изменения всех трех типов (фото 6-2). Кроме сферических вирионов, образуются нити, обычно содержащие несколько нуклеокапсидов, из которых только один необходим для инфекционности (Компанс и др., 1966; Хосака и др., 1966).
Мы перечислили морфологические изменения, ведущие к почкованию, произвольно расположив их в определенном порядке, тогда как точная последовательность событий все еще неизвестна. Многое должно обусловливаться сродством определенных вирусных белков к другим белкам и к липидам плазматической мембраны.
Мембранный белок М, по-видимому, является мишенью, в которую «целится» РНП; совершенно очевидно, что два взаимодействующих белка имеют сродство друг к другу. При одновременном заражении клеток вирусом SV5 и вирусом везикулярного стоматита (ВВС) в потомстве образуются в большом количестве фенотипически смешанные частицы, по форме напоминающие пулю. Они содержат нуклеокапсид (и, следовательно, геном) и все белки ВВС, а также два гликопротеида пепломеров SV5, но белок М вируса SV5 не входит в их состав (Мак-Шарри и др., 1971). Это согласуется с гипотезой о том, что белок М ВВС узнается только своим собственным РНП.
Нуклеокапсиды парамиксовирусов могут образовывать в цитоплазме крупные включения, четко различимые в электронном или в световом микроскопе, где они видны как большие несимметричные ацидофильные включения, характерные именно для рода Paramyxovirus. В изученных системах вирус — клетка величина избытка накапливающихся нуклеокапсидов, по-видимому, обратно пропорциональна урожаю инфекционного вируса (Компанс и др., 1966).
- Читать далее "Метаболизм пораженных парамиксовирусами клеток. Слияние клеток пораженных парамиксовирусами"
Оглавление темы "Характеристика парамиксовирусов и рабдовирусов":1. Метаболизм клетки пораженной вирусом гриппа. Зависимость вируса гриппа от клеток хозяина
2. Неполный вирус гриппа. Парамиксовирусы
3. Ранняя стадия парамиксовирусной инфекции. Транскрипция парамиксовирусов
4. Трансляция и репликация РНК парамиксовирусов. Сборка и выход парамиксовирусов из клетки
5. Метаболизм пораженных парамиксовирусами клеток. Слияние клеток пораженных парамиксовирусами
6. Размножение коронавирусов человека. Размножение аренавирусов и супергруппы Буниамвера
7. Вирус везикулярного стоматита. Цикл размножения и транскрипция рабдовирусов
8. Трансляция рабдовирусов. Репликация РНК рабдовирусов
9. Сборка и выход рабдовирусов из клетки. Степень выраженности генов рабдовирусов
10. Метаболизм пораженных рабдовирусом клеток. T-частицы рабдовирусов