Бактериальные плазмиды: функции, задачи
Плазмидами у бактерий называются кольцевые, реже линейные автономно реплицирующиеся молекулы ДНК (репликоны), без которых клетка способна существовать (в отличие от хромосом, без которых клетка существовать не может). Иногда в определенных условиях существования плазмидные функции приобретают жизненно важное значение для бактериальной популяции. Первой обнаруженной у бактерий плазмидой, как уже было сказано выше, была F-плазмида Е. coli, или половая плазмида.
Указанием на наличие плазмиды (или плазмид) в бактериальной клетке является быстрый перенос каких-то признаков от донора к реципиенту при конъюгации. Естественно, что таким способом обнаруживаются только половые плазмиды, контролирующие какие-то фенотипически проявляющиеся свойства. Указанием на присутствие плазмиды в бактериальной клетке может быть и отсутствие генетического сцепления каких-то маркеров с хромосомными генами, установленное при генетическом анализе. Этот критерий выявления плазмид можно использовать только у хорошо генетически изученных видов бактерий, к тому же автономным репликоном может быть не плазмида, а вторая хромосома. В ряде случаев можно идентифицировать плазмиды по их репликативной автономности.
Процесс репликации у плазмид обычно более чувствителен, чем у хромосом, к некоторым ингибиторам: акридиновым красителям, додецилсульфату натрия, ионам висмута и кремния, высокой температуре. При воздействии этими ингибиторами контролируемые плазмидами фенотипические признаки теряются раньше, чем свойства, обусловленные хромосомными генами. Универсальным, пригодным для всех бактерий, методом обнаружения плазмид в бактериальной клетке является выявление их при электрофоретическом анализе.
Как уже говорилось, количество и размеры плазмид в клетках разных бактерий могут сильно варьировать: крупные плазмиды (более 20 тпо) обычно представлены в бактериальной клетке одной копией, хотя типов таких плазмид у одной бактерии может быть несколько. Мелкие плазмиды (менее 20 тпо) обычно многокопийны и одна бактерия тоже может содержать несколько их типов. Таким образом, суммарное количество плазмидной ДНК в клетке иногда превосходит количество хромосомной ДНК.
Для мелких плазмид характерна способность к сверхрепликации — реплицируются не синхронно с хромосомой, а намного чаще, что и обеспечивает их многокопийность в клетке. Крупные плазмиды в норме реплицируются синхронно с хромосомой, что обеспечивается их прикреплением к клеточной мембране в области специального района их ДНК — par-локуса. Однако у крупных конъюгативных плазмид тоже наблюдается способность к сверхрепликации во время конъюгационного переноса при синтезе ДНК с ori Т-последовательности плазмиды.
Очевидно, что при общем огромном числе плазмид у бактерий и большом их разнообразии возникает необходимость в классифицикации. Группировка плазмид базируется в основном на их фенотипических признаках и не связана с филогенетическим родством. Единственным разделением, близким к истинной классификации, является компановка плазмид по группам несовместимости. Несовместимостью называется неспособность близкородственных плазмид поддерживаться (т.е. существовать) в одной клетке. В настоящее время выделено большое число групп несовместимости у плазмид различных бактерий. Ниже мы вернемся к этому свойству плазмид.
Среди признаков, по которым группируются плазмиды, не связанных с истинной классификацией, можно назвать приведенное выше разделение по размеру: крупные и мелкие. Некоторые авторы делят плазмиды на эписомные, т. е. способные встраиваться в хромосому (эписома—плазмида, интегрированная в хромосому), и неэписомные. Однако такое разделение некорректно, поскольку способность плазмиды быть эписомой зависит не только от ее особенностей, но и от структуры хромосомы: в клетках одного вида бактерий плазмида может быть эписомой, а в клетках другого вида—нет. Плазмиды делят на трансмиссивные и нетрансмиссивные, т.е. имеющие tra-систему и не имеющие ее (можно назвать их также конъюгативные и неконъюгативные).
Трансмиссивные плазмиды в подавляющем большинстве случаев (кроме плазмид актиномицетов) крупные, поскольку tra-область таких плазмид включает более двадцати генов. Нетрансмиссивные плазмиды могут быть и мелкими, и крупными. Некоторые конъюгативные плазмиды передаются только в пределах одного или близких видов, другие могут передаваться при конъюгации разным видам и даже разным родам бактерий. Такие плазмиды называются широкотрансмиссивными.
Можно разделять плазмиды по типу их репликации: одним плазмидам для репликации нужен синтез плазмидоспецифичных белков de novo, другим — не нужен. Наконец, можно компановать плазмиды в группы согласно определяемым ими фенотипическим признакам бактерий. Таких признаков очень большое количество, причем многие из них существенны не только для бактериальной клетки, но и для человека. Приведем отдельные, конечно, далеко не все, свойства бактерий, контролируемые плазмидами.
Некоторые плазмиды имеют гены, кодирующие синтез бактериоцинов — соединений, подавляющих рост других бактерий. Наиболее хорошо изученными бактериоцинами являются колицины — молекулы относительно простых белков или липогликопротеидов, абсорбирующихся на клеточных мембранах бактерий и мешающих осуществлению жизненно важных реакций в клетке. Многие плазмиды бактерий колийной группы имеют гены, контролирующие синтез колицинов и ген, определяющий резистентность к ним, так что на хозяина собственный колицин не действует. Известны и другие бактериоцины.
Очень многие плазмиды несут гены, определяющие устойчивость (резистентность) к различным токсичным для бактерий препаратам: антибиотикам, тяжелым металлам и другим повреждающим ДНК агентам. Такие плазмиды называют R-плазмидами (плазмидами резистентности).
Большая группа плазмид, встречающихся в основном у различных почвенных бактерий, имеет гены, контролирующие способность к деградации ксенобиотиков — соединений углеводородов, камфоры, толуола, хлора и других продуктов, не утилизируемых не содержащими таких плазмид бактериями. Эти плазмиды называются плазмидами деградации, или D-плазмидами.
Многие плазмиды патогенных бактерий несут гены, контролирующие их вирулентные свойства, такие как резистентность к бактерицидному действию сыворотки крови, синтез энтеротоксинов и гемолизинов, синтез поверхностных антигенов, способствующих прикреплению бактериальных клеток к эпителию кишечника. Иногда патогенный бактериальный штамм отличается от непатогенного только наличием определенной плазмиды.
Некоторые бактериальные плазмиды кодируют синтез антибиотиков, ферментов рестрикции и модификации ДНК, возможность использования различных сахаров. Иногда плазмиды несут гены, контролирующие синтез продуктов, аналогичных соединениям, образование которых зависит от хромосомных генов, например некоторых белков, участвующих в синтезе ДНК.
Плазмиды фитопатогенных бактерий могут определять их вирулентность для растений. Так, способность образовывать опухоли при заражении растений бактерией Agrobacterium tumefaciens обусловлена наличием у последней крупной плазмиды, контролирующей синтез и катаболизм опинов; способность к азотфиксации у клубеньковых бактерий (ризобий) детерминирована так называемыми симбиотическими плазмидами (Sym-плазмидами).
Существуют и плазмиды, присутствие которых в бактериальных клетках не приводит к каким-то заметным фенотипическим эффектам. Такие плазмиды называются криптическими. Однако это понятие относительно, так как часто оказывается, что в каких-то условиях указанные плазмиды обусловливают определенные фенотипические признаки содержащих их бактерий.
В принципе в состав плазмиды может быть включен любой бактериальный ген, который далее может быть передан с плазмидой другим бактериям того же либо другого вида и даже рода. Наиболее интенсивно и широко происходит передача плазмид при конъюгации. При этом способе передачи генов у бактерий (от донора к реципиенту) переносятся трансмиссивные, или конъюгативные (половые), плазмиды, имеющие в своем составе систему tra-генов (tra+). Однако такие плазмиды, обеспечив процесс конъюгации бактериальных клеток, могут мобилизовать на перенос и плазмиды, не имеющие tra-системы, если в их ДНК есть последовательность, гомологичная ori Т-последовательности трансмиссивной плазмиды и опознаваемая эндонуклеазой tra-системы.
Такая последовательность в молекулах нетрансмиссивных плазмид называется mob-сайтом (от мобилизации), а сами плазмиды—мобилизуемыми. Очень многие нетрансмиссивные бактериальные плазмиды являются мобилизуемыми (tra-mob+). Кроме передачи при конъюгации, плазмиды могут распространяться от бактерии к бактерии и за счет трансформации и трансдукции плазмидной ДНК. Отдельные плазмидные гены или группы генов могут быть переданы в процессе транспозиции (см. следующий раздел).
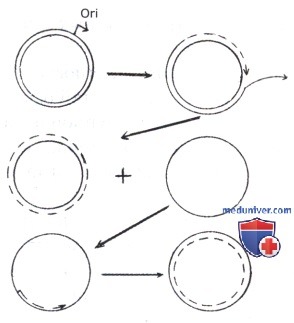
Репликация плазмид инициируется в плазмидной последовательности ori V и заканчивается на плазмидном терминаторе—ter. При этом у части плазмид терминатор находится, как у хромосомы, напротив ori V, на противоположной дуге кольцевой молекулы, и репликация идет от ori V в противоположных направлениях. У других плазмид терминатор находится вблизи ori V, и репликация идет от ориджина в одну противоположную от терминатора сторону и заканчивается на нем. У ряда плазмид обнаруживается несколько ori V. По-видимому, это связано с их гибридной природой — возникли из нескольких плазмид в результате их рекомбинации. У таких плазмид работает один ori V, а другие зарепрессированы.
Если он повреждается или вырезается, начинает работать другой ориджин. Для репликации всех плазмид нужен основной фермент репликации — ДНК-полимераза III, для репликации мелких плазмид необходима, кроме того, ДНК-полимераза I — фермент, участвующий в процессах репарации ДНК; помимо этого нужны и другие белки, кодируемые хромосомой. Ряду плазмид для репликации, как уже было сказано выше, нужен синтез плазмидоспецифичного белка de novo.
Некоторые бактериальные плазмиды, например плазмиды стафилококков, реплицируются по механизму катящегося кольца, как половые плазмиды при конъюгативном переносе. Репликация инициируется на одной нити — специфичная эндонуклеаза делает однонитевой разрез в последовательности ori V, и одна нить как бы соскальзывает с двунитевой кольцевой молекулы. Одновременно идет синтез ДНК по оставшейся кольцевой матричной нити. В результате возникают две кольцевых молекулы плазмидной ДНК: однонитевая и двунитевая. Далее на однонитевой молекуле инициируется синтез второй нити, и она превращается в двунитевую. Одноцепочечная ДНК—активный интермедиат в рекомбинационных процессах, поэтому плазмиды, реплицирующиеся по механизму катящегося кольца, часто претерпевают рекомбинацию.
С процессом репликации тесно связано такое свойство плазмид, как их несовместимость. Показано, что у несовместимых плазмид идентична структура РНК I — мелкой РНК, ингибирующей начало нового цикла репликации. По-видимому, какую-то роль в феномене несовместимости у крупных плазмид играет и возможность их прикрепления к клеточной мембране.
Кроме несовместимости близкородственных плазмид известен и другой феномен их взаимодействия — исключение вхождения, т.е. невозможность плазмиды проникнуть в бактериальную клетку, если там уже есть сходная плазмида. За этот эффект отвечают гены tra-системы плазмиды; известен специфический белок, участвующий в исключении. Феномен исключения вхождения слабо выражен в старых бактериальных культурах, так что может быть преодолен при необходимости.
В бактериальных клетках, содержащих две неидентичные плазмиды или более, возможен обмен фрагментами молекул (ДНК). При этом может происходить как контролируемый общей системой рекомбинации обмен фрагментами с перекрестами по имеющимся участкам гомологии, так и образование коинтегратов двух или большего числа плазмид с участием мигрирующих элементов (см. следующий раздел). При разрешении (распадении) таких коинтегратов возможно образование новых последовательностей ДНК. (Возможно наличие нескольких ori V в одной плазмиде — последствие таких коинтегратов.)
Кроме взаимодействия плазмид друг с другом, в клетке возможны и плазмиднохромосомные взаимодействия. Выше уже были описаны коинтеграты конъюгативных плазмид с хромосомой (возникновение донорных штаммов Hfr), разрешение которых может быть правильным и неправильным. Могут встраиваться в хромосому и неконъюгативные плазмиды: в результате процесса общей рекомбинации, если у двух этих структур есть гомологичные последовательности, или в результате транспозиции мигрирующих элементов. Кроме структурных взаимодействий, плазмидные и хромосомные гены в ряде случаев осуществляют кооперативный контроль некоторых клеточных процессов. Так, гены хромосомы участвуют в контроле процессов, зависящих от плазмиды, таких как синтез пилей, образование конъюгирующих пар клеток и т.д. Гены плазмид влияют на свойства бактерий, контролируемые хромосомными генами: жизнеспособность бактерий, уровень мутагенеза и рекомбинации, споруляцию и синтез ряда соединений.
В целом плазмиды играют весьма существенную роль не только в жизни отдельных бактериальных клеток, но и бактериальных популяций.
- Читать далее "Мигрирующие элементы ДНК бактерий: инсерционные последовательности и транспозоны"
Редактор: Искандер Милевски. Дата публикации: 14.07.2019
- Геном бактерий - строение бактериальной ДНК
- Формы переноса генетического материала бактерий: трансформация
- Трансдукция у бактериофагов и бактерий
- Конъюгация у бактерий
- Слияние протопластов бактерий
- Бактериальные плазмиды: функции, задачи
- Мигрирующие элементы ДНК бактерий: инсерционные последовательности и транспозоны