Формы переноса генетического материала бактерий: трансформация
У бактерий известно три естественных способа передачи генетического материала: генетическая трансформация, конъюгация и трансдукция. Кроме того, у них происходит передача отдельных генов или групп локализованных рядом генов за счет их перемещения путем транспозиции. (Процесс транспозиции будет рассмотрен в разделе, посвященном мигрирующим элементам бактерий.) Кроме естественных способов передачи генов, существует разработанный для бактерий искусственный метод слияния клеток (их гибридизации), названный слиянием протопластов. Мы остановимся на трех первых из упомянутых выше природных механизмов передачи генов и коротко охарактеризуем метод слияния протопластов. Необходимо иметь в виду, что все эксперименты по обмену генетической информацией между разными штаммами бактерий проводились на генетически маркированных культурах — мутантах с ярко выраженными и поддающимися учету при высевах на разные селективные среды признаками.
Трансформация — это приобретение бактерией-реципиентом свойств бактерии-донора в результате поглощения ДНК последней. Явление было открыто в 1928 г. на пневмококке (Streptococcus pneumoniae) Гриффитом (Griffith F.), который продемонстрировал превращение непатогенных R-форм в патогенные S-формы в результате совместного введения мышам живых бактериальных R-клеток и убитых S-клеток. В то время значимость обнаруженного явления не была понята, и к исследованию этого феномена вернулись только в 40-е годы. В 1944 г. Эвери (Avery G. J.) было показано, что трансформирующим фактором является ДНК. В конце 40-х годов для трансформации стали использовать высокоочищенные препараты ДНК; кроме того, трансформирующая активность ДНК была продемонстрирована в экспериментах с радиоактивной меткой.
Механизмы трансформации наиболее хорошо изучены на пневмококке (Streptococcus pneumoniae), сенной палочке (Bacillus subtilis) и гемофилле (Hemophillus influenzae).
Процесс трансформации, как и всякий перенос ДНК у бактерий, имеет две стадии: перенос ДНК в клетку-реципиент и включение ДНК донора в хромосому реципиента.
Чтобы трансформация прошла успешно, донорная ДНК должна обладать определенными свойствами. Для эффективных абсорбций ДНК на клетке-реципиенте и поглощения ее необходимо, чтобы она была двунитевой. Однонитевая ДНК практически не трансформирует реципиента (эффективность трансформации 0,5% от уровня трансформации препаратом нативной ДНК). Правда, при проведении трансформации при кислых значениях pH (в норме наибольшая активность процесса достигается при нейтральном pH) эффективность трансформации однонитевой ДНК увеличивается. Кроме того, у гемофилов есть мутанты, которые трансформируются только однонитевой ДНК. Помимо двунитчатости трансформирующие фрагменты ДНК должны иметь определенный размер: их протяженность должна превышать 450 по (0,3 MD). Обычно получают препараты ДНК с размерами фрагментов 22-45 тпо (15-30 MD), что составляет 1-2% длины хромосомы. Такие фрагменты эффективно трансформируют бактериальные клетки. Если увеличить длину фрагментов ДНК, выделяя их мягкими методами, это не сказывается на эффективности трансформации, в ее процессе они все равно фрагментируются. У большинства бактерий при трансформации поглощается любая (имеются в виду ее последовательности) ДНК; у гонококков и гемофилов для успешного поглощения во фрагментах ДНК должны присутствовать определенные последовательности.
Кроме перечисленных выше свойств фрагментов донорной ДНК, для успешной трансформации необходимо определенное состояние клеток реципиента— компетентность (способность абсорбировать и поглощать ДНК донора). Большинство, но не все бактерии находятся в состоянии компетентности в конце фазы логарифмического роста. При этом у ряда бактерий, таких как пневмококки, стрептококки, гонококки, гемофилы могут быть компетентны все клетки культуры, у других, например у сенной палочки,—только некоторые клетки в культуре.
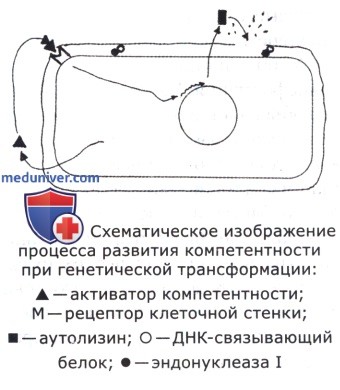
Компетентные клетки характеризуются сниженным метаболизмом, измененной клеточной поверхностью, повышенной чувствительностью к осмотическому шоку. Состояние компетентности обусловлено выработкой бактериальными клетками низкомолекулярного белка—фактора компетентности. Этот белок взаимодействует со специфичными рецепторами клеточной стенки и активирует синтез других белков компетентности: аутолизина, ДНК-связывающего белка и эндонуклеазы I. Аутолизин разрушает структуру наружной клеточной стенки бактериальных клеток и обнажает ДНК-связывающий белок, а также эндонуклеазу I. Такая клетка готова к абсорбции двунитевых фрагментов ДНК.
Эффективность трансформации зависит от концентрации ДНК до какого-то предела. Сначала эта зависимость экспоненциальна, далее характеризующая этот процесс кривая выходит на плато, что указывает на ограниченное число воспринимающих ДНК рецепторов на поверхности компетентных клеток. О том же свидетельствует уменьшение частоты трансформации при использовании смеси гомологичной и негомологичной ДНК (поглощается компетентными клетками любая ДНК, а трансформирует только гомологичная, отсюда снижение частоты трансформации). У различных видов бактерий число рецепторов на поверхности компетентных клеток довольно сильно различается; например, у сенной палочки — 10-40, у пневмококка—30-80. Кроме того, у разных авторов число рецепторов на поверхности одних и тех же бактерий не совпадает.
Абсорбция ДНК идет на всех бактериальных клетках, компетентных и некомпетентных, однако в первом случае она необратима, а во втором—обратима. Рецепторами для фрагментов ДНК являются молекулы ДНК-связывающего белка. Они абсорбируют фрагменты донорной ДНК размерами 15-30 MD (22-45 тпо). Эндонуклеаза I делает на этих фрагментах одноцепочечные разрывы на расстоянии 6 тпо друг от друга.
В клетку фрагменты входят одним концом.
Во время вхождения ДНК фрагментируется до отрезков в 2-4 тпо, и одна из цепей деградирует. Однонитевые фрагменты связываются с белком, который защищает их от деградации.
Общее количество поглощенной одной клеткой ДНК у разных авторов сильно различается — от одного до многих геномов. Однонитевые фрагменты ДНК донора входят в синапс с молекулой ДНК реципиента (хромосомой), удерживаются они водородными связями. Далее нить донора включается в молекулу ДНК реципиента, вытесняя одну из ее цепочек. Образуется гетеродуплексный участок молекулы ДНК реципиента, а лишний ее однонитевой участок элиминируется. Встраивание в хромосому реципиента одной нити донорной ДНК было продемонстрировано в генетических экспериментах. Протяженность встроившегося фрагмента обычно составляет около 3,5 тпо (2 MD), т.е. 0,1-0,2% хромосомы. Включение фрагмента донорной ДНК в хромосому реципиента происходит очень быстро—за несколько минут, для фенотипического его проявления необходимы 1-2 ч.
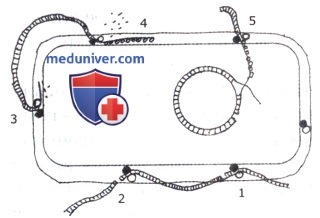
1 —абсорбция ДНК связывающим белком и одноцепочечные разрывы;
2 — расщепление ДНК на фрагменты;
3 — акцепция одной цепи ДНК и деградация другой;
4 — связывание одноцепочечной ДНК белком;
5 — интеграция трансформирующей ДНК в хромосому реципиента.
В настоящее время генетическая трансформация обнаружена у многих видов бактерий, включая актиномицеты и цианобактерии. В тех случаях, когда в стандартных условиях ее не удается получить, как у Е. coli, например, применяют специальные среды (так называемая кальцевая среда) и специальные мутантные формы бактерий. Барьер проникнования фрагментов ДНК в клетку трансформируемой бактерии можно преодолеть посредством электропорации (индукции пор в оболочках клеток-реципиентов электрическим разрядом) или использованием не интактных клеток реципиента, а их протопластов. Другими словами, сегодня препятствий к осуществлению генетической трансформации на любой бактериальной культуре не существует.
У большинства бактерий, кроме внутривидовой, возможна и межвидовая трансформация. Частоты межвидовой трансформации ниже, чем внутривидовой, что объясняется неполной гомологией геномов донора и реципиента.
Механизмы генетической трансформации у разных видов в основном одинаковы. У гемофилов отличается от описанного выше процесса первый этап трансформации — детали проникновения в клетку-реципиент фрагментов донорной ДНК. У этих бактерий на клеточной оболочке в состоянии компетентности образуются специальные выпячивания цитоплазматической мембраны — трансформасомы. На этих цитоплазматических пузырьках локализованы молекулы белка, связывающие ДНК донора. При этом связываются только фрагменты ДНК, имеющие в своем составе определенные последовательности из одиннадцати нуклеотидов. Далее процесс трансформации протекает так же, как у других бактерий, т.е. как описано выше.
Трансформировать бактериальные клетки может не только хромосомная ДНК, но и плазмидная, а также фаговая. Генетическая трансформация широко используется для введения в бактериальные клетки нужных плазмид, в том числе векторных (полученных генно-инженерным путем). Процесс генетической трансформации бактериальной клетки бактериофагами называется трансфекцией и используется для введения в бактерии фагов, которые не инфицируют их естественным путем. При трансфекции фаговая ДНК фрагментируется и часть ее, по-видимому, распадается: показано, что для образования жизнеспособной фаговой частицы при трансфекции у сенной палочки реципиентная бактериальная клетка должна поглотить 4-5 геномов фага.
Закономерности генетической трансформации, описанные выше, изучены в лабораторных экспериментах. Но несомненно, процесс этот происходит и в природных условиях. У ряда бактерий описано явление спонтанной трансформации, связанное с выходом из клетки ДНК при автолизе в старых культурах: у сенной палочки обнаружен выход ДНК из клетки как упорядоченный процесс. У пневмококков показана спонтанная генетическая трансформация в организме зараженных животных. Безусловно, генетическая трансформация у бактерий осуществляется в природных условиях при больших плотностях их популяций, особенно, по-видимому, в бактериальных пленках и скоплениях в различных экологических нишах, включая организмы растений, животных и человека.
- Читать далее "Трансдукция у бактериофагов и бактерий"
Редактор: Искандер Милевски. Дата публикации: 12.07.2019
- Геном бактерий - строение бактериальной ДНК
- Формы переноса генетического материала бактерий: трансформация
- Трансдукция у бактериофагов и бактерий
- Конъюгация у бактерий
- Слияние протопластов бактерий
- Бактериальные плазмиды: функции, задачи
- Мигрирующие элементы ДНК бактерий: инсерционные последовательности и транспозоны