Геном бактерий - строение бактериальной ДНК
Генетика бактерий включает очень широкий круг вопросов. Это строение и организация генома, изменчивость и мутагенез, формы переноса генетического материала и генетический анализ, генетический контроль различных жизненно важных процессов, таких как репликация, рекомбинация, репарация, транскрипция и трансляция, регуляция генного действия, генетические основы селекции. Охватить их в одной статье невозможно. Наиболее важны для медицинской микробиологии правильные представления об организации генетического материала бактерий, формах его переноса и влиянии этого переноса на изменчивость бактерий. Этим вопросам и посвящены последующие статьи на сайте.
В аспекте медицинской микробиологии важны также сведения о мутационном процессе у бактерий. Данные об участии в нем перемещающихся элементов, вносящих основной вклад в процессы изменчивости бактерий, приводятся в отдельных статьях на сайте. Классические материалы о классификации мутаций и их индукции химическими и физическими факторами присутствуют во всех учебниках по генетике и в эту статью не включены.
Генетическим материалом у всех живых организмов являются нуклеиновые кислоты. В подавляющем большинстве случаев, в том числе и у бактерий, это дезоксирибонуклеиновая кислота (ДНК); у некоторых вирусов как исключение эту роль выполняет рибонуклеиновая кислота (РНК). ДНК представляет собой макромолекулу, образованную повторяющимися структурами — нуклеотидами. Каждый нуклеотид состоит из трех компонентов: циклического азотсодержащего соединения, называемого основанием, сахара пентозы и фосфорной кислоты. В ДНК присутствуют четыре главных азотистых основания: два пиримидиновых — тимин и цитозин и два пуриновых — аденин и гуанин (в РНК тимин заменен урацилом). Наряду с главными основаниями у некоторых организмов в ДНК присутствуют минорные редко встречающиеся основания: 5-метилцитозин, 5-оксиметилцитозин и др.
Сахар пентоза в составе ДНК—2-дезоксирибоза (отсюда дезоксирибонуклеиновая). Часть нуклеотида, состоящая из сахара с присоединенным к нему азотистым основанием, называется нуклеозидом. Поэтому нуклеотиды называют и нуклеозидмонофосфатами.
В любой молекуле ДНК сумма нуклеотидов с пуриновыми основаниями равна сумме нуклеотидов с пиримидиновыми основаниями, т. е. А + Г = Т + Ц. Содержание аденина (А) равно содержанию тимина (Т), а гуанина (Г) — содержанию цитозина (Ц); таким образом, отношения А к Т и Г к Ц равны единице. Кроме того, Г + Т = А + Ц. Отношение А + Т к Г + Ц видоспецифично и может значительно варьировать в ДНК организмов разных видов.
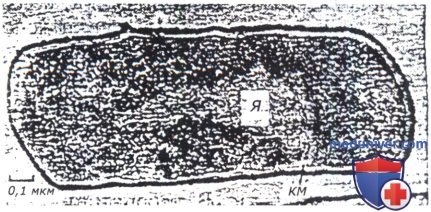
В целом ДНК представляет собой молекулу, содержащую две полинуклеотидные цепи. Эти цепи закручены вокруг общей оси в двойную спираль, витки которой, если смотреть вдоль оси спирали, идут по часовой стрелке. Такая конфигурация ДНК называется правозакрученной и обозначается как В-форма. В живых клетках большинство молекул В-ДНК претерпевает дополнительную спирализацию, что облегчает процессы репликации (удвоения) и рекомбинации (обмена фрагментами)Цепи ДНК образованы последовательностью нуклеотидов, соединенных фосфодиэфирными связями, объединяющими две 2-дезоксирибозы, к которым присоеденены азотистые основания. Спиральная конфигурация молекулы ДНК поддерживается водородными связями между основаниями двух цепей. В результате спаренные основания располагаются между двумя цепями перпендикулярно их оси. Структура в целом похожа на винтовую лестницу, каркас которой образован сахарофосфатными цепями молекулы, а ступеньками служат спаренные основания.
Существенным моментом в структуре двойной спирали ДНК является принцип комплементарности: спаривание оснований происходит строго специфично — аденин всегда соединен с тимином, а цитозин — с гуанином, т. е. каждая пара оснований содержит по одному пурину и одному пиримидину.
Молекула ДНК выполняет две функции: аутокаталитическую (способность к репликации) и гетерокаталитическую (управление экспрессией генов). При репликации ДНК ее цепи частично раскручиваются и разделяются в результате разрыва водородных связей между комплементарными основаниями. Образовавшиеся одноцепочечные участки исходной молекулы служат матрицей, к которой по принципу комплементарности оснований присоединяются соответствующие нуклеотиды. Последние соединяются фосфодиэфирными связями, образуя новую цепь, комплементарную родительской. В результате из одной молекулы образуются идентичные ей две двухцепочечные структуры. Такая репликация называется полуконсервативной: каждая из вновь образовавшихся молекул состоит из одной старой (родительской) цепи и одной новой (дочерней). В процессе репликации участвует целый комплекс ферментов (см. следующий раздел).
Гетерокаталитическая функция ДНК осуществляется в ходе процессов транскрипции — синтеза одной цепи информационной, или матричной, РНК— иРНК (по матрице ДНК) и трансляции этой иРНК на рибосомах—сборке аминокислот в полипептидную цепь (по матрице иРНК).
Четкость гетерокаталитической функции ДНК обеспечивается генетическим кодом, записанным в молекуле ДНК. Единица генетической информации, определяющая, какая из аминокислот будет встраиваться в полипептид (молекулу белка), называется кодоном. Кодон состоит из трех нуклеотидов. Другими словами, генетический код триплетен. Он не перекрывается, т.е. соседние триплеты не имеют общих оснований, и считывается без запятых, т.е. не содержит разделительных знаков между триплетами. Код колинеарен. Это означает, что порядок расположения кодонов в ДНК (и следовательно, в иРНК) совпадает с порядком расположения аминокислот в кодируемом белке. Генетический код универсален для всех живых организмов. Правильную трансляцию генетической информации во время синтеза белка обеспечивают инициирующие и терминирующие кодоны, определяющие начало и конец синтеза генного продукта—белка. Кроме последовательностей, кодирующих белки, в ДНК имеются специфические регуляторные области, опознаваемые регуляторными белками.
Несмотря на большое разнообразие бактерий, внутренняя структура всех бактериальных клеток и их генетический аппарат (ДНК) практически одинаковы. С точки зрения генетики лучше всего изучены палочковидные истинные бактерии — кишечная палочка (Escherichia coli) и сенная палочка (Bacillus subtilis). Геном всех бактерий включает ядерный аппарат — «хромосому», без которой клетка существовать не может, и плазмиды — молекулы ДНК, без которых бактериальная клетка может выжить (они жизненно необходимы только в определенных условиях существования).
Ядерный аппарат бактерий называют нуклеоидом: от истинного клеточного ядра он отличается отсутствием ядерной мембраны и тем, что при делении клетки не происходит его реорганизация. Нуклеоид связан с клеточной мембраной посредством выроста последней — органеллы, называемой мезосомой. Фактически нуклеоид представляет собой молекулу ДНК. У подавляющего большинства бактерий эта молекула кольцевая, хотя обнаружены бактерии, у которых она имеет линейную форму. В функциональном смысле нуклеоиды бактерий являются настоящими хромосомами, хотя структурная организация их более проста, чем структура хромосом эукариотов.
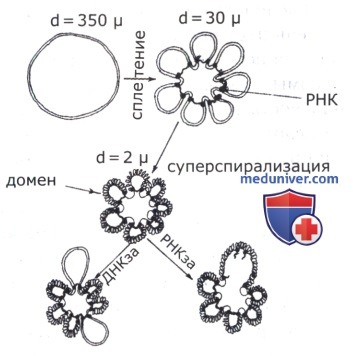
Впервые кольцевое строение хромосомы кишечной палочки прямо было показано в 1963 г. Кернсом (Cairns J.), получившим ее электронно-микроскопическую фотографию. (До этого существовали генетические данные, на основании которых был сделан такой же вывод.) Длина молекулы ДНК, представляющей собой бактериальную хромосому, по его расчетам, составляла приблизительно 1 мм. Первая модель упаковки кольцевой молекулы ДНК в нуклеоиде была предложена Петтиджоном (Pettijohn D. F.) в 1974 г., и до сих пор принципиально в ней мало что изменилось. По этой модели, ДНК находится в суперскрученном состоянии, что обусловлено ферментом топоизомеразой.
Скрученная нить собрана в крупные петли — домены, удерживаемые молекулами РНК. В результате диаметр нуклеоида составляет 1 мкм при длине хромосомы 1,6 мм. Сейчас показано, что молекулы РНК, присутствующие в нуклеоиде, — это информационная (иРНК) и транспортная (тРНК) РНК. В 1995 г. были опубликованы материалы, неопровержимо доказывающие наличие в нуклеоиде около 20 различных мелких белков, которые, по-видимому, участвуют в организации структуры нуклеоида— укладке молекулы ДНК. Было показано также, что на клеточной мембране у бактерий, помимо мезосомы, есть и другие участки прикрепления к ней ДНК: у Е. coli их 20-30, у Вас. subtilis — 70-90. Возможно, именно участки прикрепления молекулы ДНК к клеточной мембране играют определенную роль в организации доменной структуры нуклеоида. В 1994 г. Келленбергером (Kellenberger G.) была предложена модель нуклеоида в виде кочана цветной капусты с выступающими из него петлями—доменами.
Репликация бактериальной хромосомы занимает весь клеточный цикл, т.е. происходит постоянно. Она начинается с участка ДНК, называемого ori С (от oridgin — начало), идет в двух противоположных направлениях и заканчивается напротив ori С в районе ter С (от terminus — конец). Участок ori С у Е. coli — это последовательность ДНК протяженностью около 460 пар оснований (по), содержащая повторяющиеся фрагменты из 15 по. Ter С тоже содержит повторяющиеся последовательности, но здесь их меньше, чем в ori С. Процесс репликации осуществляется ферментными комплексами. Начинает его инициосома, состоящая из ключевого фермента, связывающегося с повторами ДНК в ori С, и геликазы, расплетающей ДНК с образованием вилки репликации. Далее работает второй комплекс — реплисома, содержащий кроме ферментов инициосомы еще ДНК полимеразу III (ДНК pol III), топоизомеразу и др. Ori С и ter С связаны с мезосомой, и эта связь играет не только структурную, но и функциональную роль, участвуя в регуляции репликации.
Весь цикл репликации у Е. coli занимает 30-40 мин. Начало каждого нового тура репликации регулируется белками цитоплазматической мембраны, к которым относится и инициосома. Новый цикл репликации обычно начинается до окончания предыдущего; иногда в ori С инициируется не только второй, но и третий, и четвертый циклы репликации до окончания первого ее тура. В связи с этим гены, расположенные вблизи от ori С, представлены в нуклеоиде большим числом копий, чем удаленные от района инициации репликации.
Расположение генов на бактериальной хромосоме неравномерно: районы с очень плотным их расположением чередуются с более разреженными участками. Эта неравномерность трактуется как результат организации (упаковки) хромосомы: разная доступность для транскрипции в разных районах. Наибольшая плотность генов регистрируется в области начала репликации. При этом здесь сосредоточены наиболее жизненно важные гены, связанные с процессами репликации, транскрипции и трансляции, рекомбинации и репарации генетического материала. Поскольку этот район, как сказано выше, мультикопиен, упомянутые гены защищены надежнее, чем расположенные дальше от ori С гены, связанные с менее важными процессами метаболизма. Наименьшая плотность генов отмечена в районе ter С. Обусловлено это тем, что данная область хромосомы насыщена сайтами (участками) рекомбинации и поэтому здесь часто возникают различные перестройки генетического материала, повреждающие гены. Отмеченный выше порядок расположения генов—тенденция, а не строгая закономерность: например, гены различных путей метаболизма встречаются во всех областях хромосомы.
Кроме хромосомы, большинство бактериальных клеток содержит плазмиды— молекулы ДНК, присутствие которых для клетки не строго обязательно. В данном разделе даются только минимальные сведения о плазмидах, основной материал об этих клеточных органеллах приведен ниже. Некоторые виды содержат только один тип плазмид, другие — по два-три типа; встречаются виды (особенно среди морских бактерий), обладающие семью — восемью различными плазмидами. Крупные плазмиды обычно имеют в клетке по одной копии, как и хромосома, связаны с клеточной мембраной и реплицируются синхронно с хромосомой. Мелкие плазмиды могут иметь в клетке много копий, реплицируются с большей интенсивностью, чем хромосома и крупные плазмиды, и распределяются при делении клетки случайно. У подавляющего большинства бактерий плазмиды, как и хромосомы, — кольцевые молекулы ДНК.
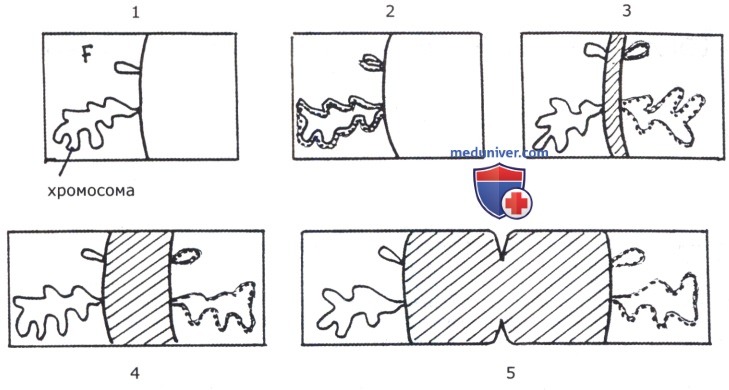
Распределение реплицирующихся в бактериях хромосом и крупных плазмид по дочерним клеткам при делении должно регулироваться, поскольку они обычно однокопийны. Первая схема расхождения этих генетических структур при делении была предложена в 1963 г. Жакобом, Бренером и Кузин (Jacob Е, Brenner S., Kusen). Согласно этой схеме, процесс расхождения пассивен. И хромосома, и крупные плазмиды прикреплены к клеточной мембране; предполагалось, что это прикрепление происходит как бы по «экватору» клетки, как раз в.той области, где происходит разрастание клеточной оболочки при увеличении объема клетки во время цитокинеза. В результате две копии генетических структур после их репликации «растаскиваются» разрастающейся клеточной стенкой в дочерние клетки. В 90-е годы на основании новых экспериментальных материалов сформировалось представление о расхождении копий генетических структур при делении клетки как об активном и упорядоченном процессе. Было показано, что в нем участвуют близкие к миозинам белки, располагающиеся на дистальных полюсах расходящихся нуклеоидов.
Последние как бы растаскиваются ими в разные стороны (в разные клетки). Мутации по генам этих полярных белков приводят к образованию безъядерных и филаментозных клеток.
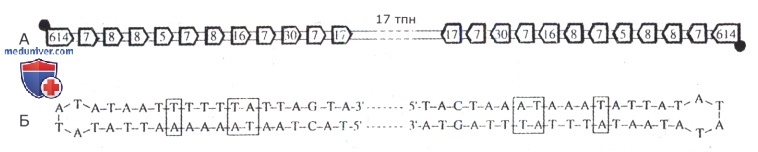
А. На обоих концах хромосомы расположены симметричные терминальные участки размером в 614 по, от них к центру идут области ДНК примерно в 800 по с «вкраплеными» в них симметричными инвертированными группами последовательностей, число которых приведено в рамках; • —белки, ковалентно связанные с 5'-концами терминальных участков хромосомы.
Б. Нуклеотидный состав инвертированных концевых участков хромосомы из 19 по, заканчивающихся «шпильками»; рамками обведены пары нуклеотидов с нарушенной симметрией.
Все сказанное выше характерно для большинства видов бактерий. Однако в самом конце 80-х годов XX века было обнаружено, что некоторые виды бактерий имеют линейные хромосомы и линейные плазмиды. Это открытие было сделано благодаря введению в круг объектов генетики новых бактериальных моделей и усовершенствованию методов исследования бактериального генома. Первая линейная хромосома была описана в 1989 г. у спирохеты Borrelia burgdorferi — возбудителя болезни Лайма (клещевой спирохетоз). Позднее линейная форма хромосом была обнаружена у различных клинических изолятов этой спирохеты. У других спирохет — лептоспир и трипонем — хромосомы кольцевые. Вскоре линейные хромосомы были выявлены и у нескольких видов актиномицетов, включая наиболее хорошо генетически изученные S. coelicolor и S. lividans.
Какова же организация линейных хромосом? В настоящее время известно два типа их организации: тип актиномицетов и тип спирохет — боррелий. У актиномицетов на концах линейных хромосом имеются молекулы белка, при помощи которых происходит превращение линейной молекулы в кольцевую. Кроме того, при анализе последовательностей ДНК (секвенировании) линейных хромосом актиномицетов было установлено, что их концы протяженностью в 25 тыс. пар оснований (тпо) состоят из инвертированных повторов — структур, по которым легко происходит рекомбинация. В результате такой рекомбинации линейные хромосомы актиномицетов могут превращаться в кольцевые. Таким образом, очевидно, существует два механизма, за счет которых линейные хромосомы актиномицетов могут преобразовываться в кольцевые: в результате рекомбинации по концевым последовательностям может возникать стабильная кольцевая структура, за счет замыкания молекулами белка — менее стабильная кольцевая структура.
Эти особенности линейных хромосом актиномицетов объясняют противоречие, возникшее при обнаружении их линейного строения; ранее существовали данные, полученные при генетическом картировании у актиномицетов, свидетельствующие о кольцевом строении их хромосом. Очевидно, хромосомы актиномицетов могут находиться в их клетках, как в линейном, так и в кольцевом состоянии. В середине линейных хромосом актиномицетов обнаружена область с последовательностью нуклеотидов, близкой к последовательности ori С Е. coli. По-видимому, репликация этих хромосом идет от центра к концам в двух направлениях. Предполагают, что так она осуществляется в кольцевой форме хромосомы, а у линейных форм репликация идет от концов к центру. У спирохет — боррелий терминальные участки линейных хромосом заканчиваются шпилечными структурами. Область ori С расположена в центре хромосомы. Вокруг нее, как и у кольцевых хромосом, находятся гены, связанные с репликацией, и гены рибосомной РНК и рибосомных белков.
У видов бактерий с линейными хромосомами обнаруживаются и линейные плазмиды. Как и у хромосом, описано два типа линейных плазмид; структуры ДНК, оканчивающиеся молекулами белка или шпильками. Тины линейных структур хромосомы и плазмид у одной и той же бактерии обычно совпадают.
Говоря о бактериальной хромосоме, мы подразумевали единичную структуру в клетке. На самом же деле бактериальные клетки часто находятся в полиплоидном состоянии — содержат несколько гомологичных ядер. Для актиномицетов и некоторых других бактерий, например, цианобактерий, — это норма. В других случаях, весьма многочисленных, когда считается, что бактериальная клетка имеет один нуклеоид, у молодых культур часто обнаруживаются клетки с двумя и даже четырьмя нуклеоидами. Кроме того, выявлены виды, имеющие по два и даже по три негомологичных нуклеоида. По два кольцевых нуклеоида имеют бруцеллы, фототрофные бактерии Rhodobacter sphaeroides, возбудитель желтухи Leptosspira interrogans. Три кольцевые хромосомы обнаружены у Pseudomonas cepacia. Фитопатогенная бактерия Agrobacterium tumefaciens имеет две хромосомы — кольцевую и линейную. Обычно у видов с несколькими негомологичными нуклеоидами одна хромосома более крупная, другая (или другие) — более мелкие.
В заключение раздела о геноме бактерий надо сказать несколько слов о полученных недавно (после 2000 г.) материалах сравнительного анализа последовательности нуклеотидов у видов с полностью секвенированным геномом. Протяженность хромосомы Е. coli составляет около 3,8 106по, средняя величина гена— 1 тпо с разбросом от 18 по (у генов половых гормонов — феромонов) до 7 тпо. Большая часть бактериальных последовательностей ДНК (85%) — структурные гены, т.е. кодирующие области. Они сгруппированы в опероны — единицы транскрипции и кластеры (группы генов, относящихся к одному метаболическому пути). Регуляторные области малы и расположены в основном между генами. Некодирующие, в основном повторяющиеся, последовательности составляют около 10% генома. Это мигрирующие элементы (см. ниже), сайты рекомбинации, регуляторы транскрипции, у гемофилов — сайты, связывающиеся с трансформасомами (см. раздел о трансформации генов). Кроме повторяющихся, к некодирующим последовательностям относятся поврежденные умеренные фаги и так называемые серые дыры — вероятно, совсем испорченные гены.
В различных бактериальных геномах встречаются и дупликации генов или очень похожие гены. Если такие гены обнаруживаются в одном и том же геноме, их называют паралогами, если в разных—ортологами. Не ясно, что представляют собой ортологи: то ли это гомологичные гены, попавшие в клетку в результате горизонтального переноса в разные виды бактерий, то ли результат конвергенции.
При сравнении прочитанных геномов разных видов был выявлен средний минимальный набор бактериальных генов, т.е. генов, контролирующих синтез нуклеотидов, репликацию, трансляцию, транскрипцию и репарацию; он подсчитан как 256 генов. Минимальный набор генов был определен и экспериментально у Bacillus subtilis. У этой бактерии инактивировали гены и учитывали те, при порче которых терялась жизнеспособность. Оказалось, что это 250-300 генов.
При анализе отсеквенированных к настоящему времени геномов оказалось, что функции приблизительно трети генов в геномах не установлены. Предполагается, что эти неизвестные гены, возможно, не функционируют в лабораторных условиях и экспрессируются только в определенных природных условиях или в каких-то природных бактериальных сообществах.
- Читать далее "Формы переноса генетического материала бактерий: трансформация"
Редактор: Искандер Милевски. Дата публикации: 12.07.2019
- Геном бактерий - строение бактериальной ДНК
- Формы переноса генетического материала бактерий: трансформация
- Трансдукция у бактериофагов и бактерий
- Конъюгация у бактерий
- Слияние протопластов бактерий
- Бактериальные плазмиды: функции, задачи
- Мигрирующие элементы ДНК бактерий: инсерционные последовательности и транспозоны