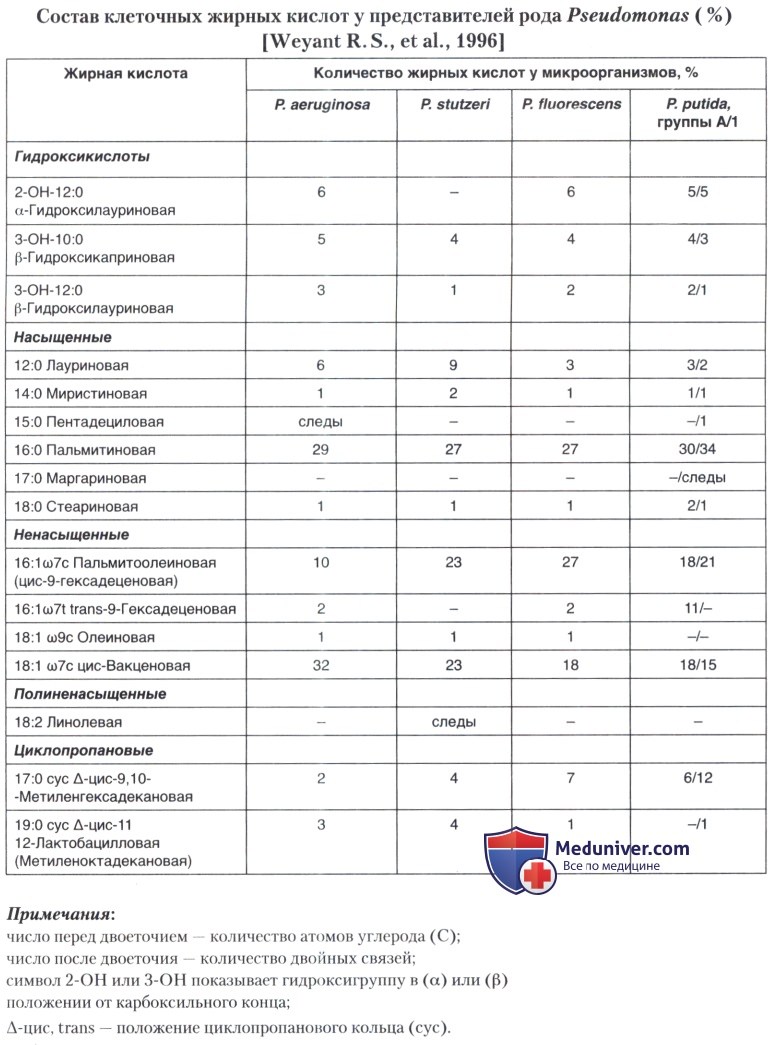
Pseudomonas stutzeri: морфология, культуральные, биохимические свойства
а) Таксономическое положение. Вид Pseudomonas stutzeri впервые описан в 1895 г. R. Burri и A. Stutzer. В семи переизданиях атласа и учебника по основам бактериологии и бактериологической диагностике (1896-1927) под редакцией К. В. Lehman и др. указаны фенотипические особенности этого микроорганизма и его таксономическое положение как Bacillus stutzeri. В 1952 г. в журнале «Journal of Bacteriology» о микробе было сообщение С. В. Van Niel и М.В. Allen. Затем в 1970-1984 гг. N.J. Palleroni и др. подтвердили таксономический статус В. stutzeri в группе аэробных псевдомонад (sensu stricto), изучая гомологию нуклеотидных последовательностей при гибридизации ДНК-ДНК и последовательностей ДНК-рРНК типового штамма и других изолятов.
В настоящее время Р. stutzeri признан принадлежащим к классу у-Proteobacteria, сем. Pseudomonadaceae, роду Pseudomonas. Филогенетическое изучение штаммов Р. stutzeri с определением последовательностей 16 S рРНК и других маркеров показало, что микроорганизм принадлежит к той же самой ветви, что Р. mendocma, Р. alcaligenes, Р. pseudoalcaligenes, Р. balearica.
б) Морфологические и физиологические признаки микроорганизма. Типичные клетки Р. stutzeri — грамотрицательные палочки (1 -3 мкм) х 0,5 мкм с одним полярным жгутиком. В некоторых случаях образуются 1-2 коротких боковых жгутика, которые чаще формируются после выращивания культуры на плотных питательных средах и легко утрачиваются при окраске. Микроорганизм имеет строго дыхательный метаболизм, образует оксидазу и каталазу, относится к денитрификаторам. P. stutzeri утилизирует мальтозу и крахмал, не гидролизует гликоген, не синтезирует аргинин-дигидролазу. По уреазной активности отмечена вариабельность.
Колонии Р. stutzeri имеют необычный вид и консистенцию. Свежеизолированные колонии напоминают кратеры вулканов. Сморщенные, кораллоиодобные, более мукоидные по периферии, непигментированные или красновато-коричневые колонии, легко снимаются с питательной среды. Облик колоний меняется со временем. При пересевах на лабораторных средах окрашенные колонии могут становиться более бледными, гладкими, маслянистыми. Темно-коричневая окраска колоний обусловлена высокой концентрацией в клетках цитохрома с.
Среди штаммов Р. stutzeri отмечена диссоциация. В 1999 г. Д.Ю. Сорокин и др. детально описали морфологию колоний R- и S-типа. Колонии R-типа продуцируют морфологически стабильные варианты, а культуры S-типа в определенных условиях образуют колонии обоих типов. Гладкие колонии, выращенные на агаре при 30°С и сохранявшиеся при 4°С в течение 24 час, часто приобретают сморщенный вид (рис. 1.9.3 — см. цветную вклейку).
Обширное исследование по использованию источников углерода для питания Р. stutzeri было выполнено R. Y. Stanier et al. в 1966 г., а затем другими учеными в 1990-х гг. с использованием как рутинных методов, так и современных тест-систем. Внутривидовая гетерогенность по 327 биохимическим признакам при утилизации углеродосодержащих соединений среди культур Р. stutzeri была очень велика. В итоге, для разграничения штаммов Р. stutzeri были выбраны 48 наиболее информативных фенотипических тестов.
При этом оказалось невозможным идентифицировать отдельные геномные группы на основе биохимических тестов. Все изученные штаммы Р. stutzeri оказались положительными, если использовались следующие субстраты: глюконат, d-глюкоза, d-мальтоза, крахмал, глицерин, ацетат, бутират, изобутират, изовалерат, пропионат, фумарат, глумарат, гликолат, глиоксилат, dl-3-гидроксибутират, итаконат, dl-лактат, dl-малат, малонат, оксалоацетат, 2-оксоглютарат, пируват, сукцинат, цитрат, d-аланин, d-аспарагин, l-глютамат, l-глютамин, l-изолейцин, l-пролин, гидролизат l-аланина-пара-нитроанилида.
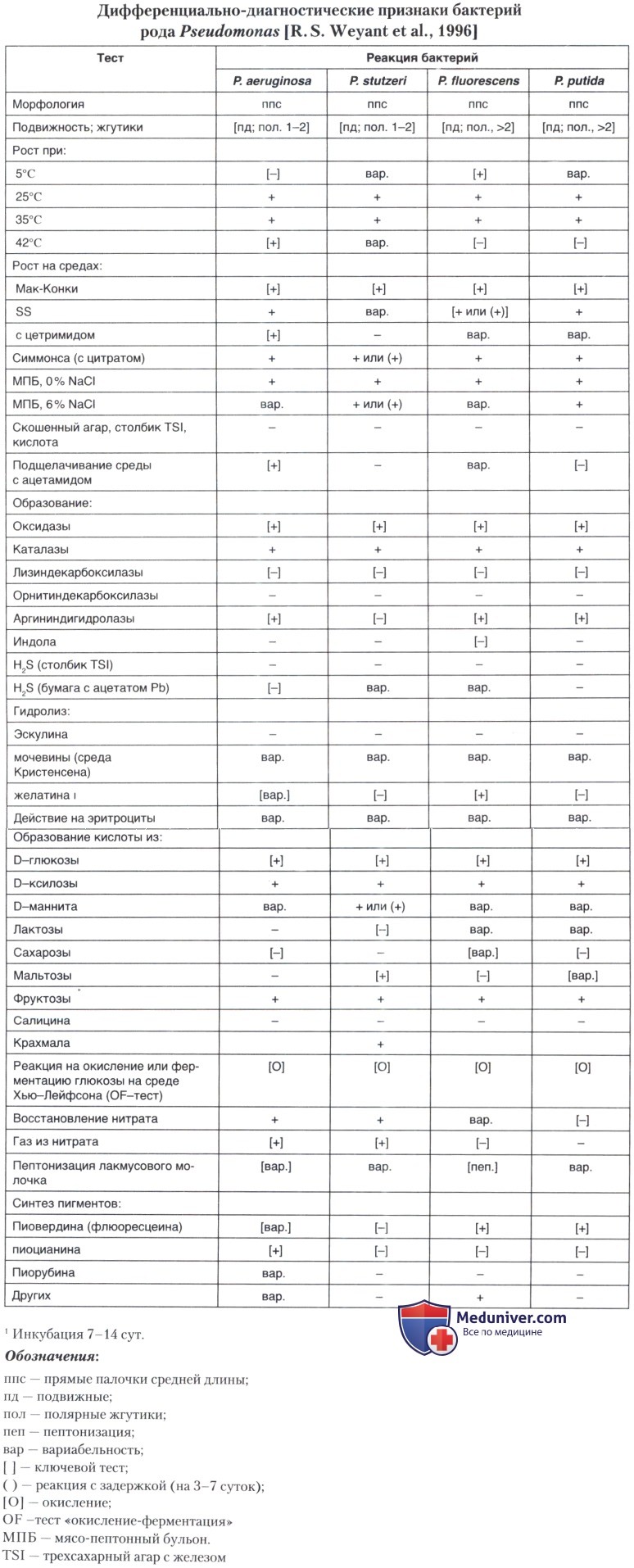
Культуры Р. stutzeri имеют широкий температурный диапазон роста. Для одних штаммов рост отмечен при температурах от 4°С до 45°С, для других температурный максимум достигает 45°С, но для большинства культур пределом является 40-41°С. Оптимальная температура для роста — около 35°С. Исследователи разделили Р. stutzeri по температурному максимуму на 2 биотипа, различающихся также содержанием гуанина (G) и цитозина (С). Штаммы первого биотипа (62 мол. % G+C) растут при температурах до 43°С. Ко второму (65-66 мол. % G+C) относятся штаммы с максимальной температурной характеристикой роста свыше 43°С.
Среди выделенных штаммов Р. stutzeri встречаются баротолерантные (выдерживают давление около 100 МПа) и барофильные (растут при гидростатическом давлении не выше 30-60 мПа). Как правило, рост при высоком давлении характерен для морских культур этого вида.
Денитрификация является стабильным физиологическим признаком Р. stutzeri. Этот гетеротрофный микроорганизм считают эталонной моделью при изучении процесса денитрификации. При изучении структур и механизмов денитрификации в одной бактерии расшифрованы примерно 50 генов, локализованных на хромосоме. Полный процесс денитрификации приводит к газообразованию N, Организация генетического кодирования дыхательной редукции нитрата у Р. stutzeri почти полностью расшифрована (nar-гены). Некоторые из nar-генов гомологичны генам нитратного дыхания у Е. coli. Они находятся у бактерии в отдельном локусе кластера nos-nis-nor. Однако точное количество генов все-таки еще не определено и требует дальнейшего изучения.
Известно, что циркулирующие в природе Р. stutzeri окисляют тиосульфат до тетратионата, используя нитриты, нитраты или N2O как электронные акцепторы. Оксидация тиосульфата в анаэробных условиях проходит медленнее, чем в присутствии O2.
Никаких флюоресцирующих пигментов бактерия не продуцирует, что позволяет отличать ее от представителей флюоресцентной группы Pseudomonas spp.
При отсутствии синтеза пигментов пиовердинов, имеющих важное физиологическое и таксономическое значение, у некоторых штаммов Р. stutzeri в условиях дефицита железа образуются катехол-подобные сидерофоры. Сидеротипирование расценивается исследователями как успешная техника для разграничения видов в пределах рода Pseudomonas. Отмечено также, что внутривидовая гетерогенность нефлюоресцирующих видов Р. stutzeri сопряжена с продукцией сидерофоров.
в) Устойчивость к внешним факторам, антисептическим и антибактериальным препаратам. Устойчивость Р. stutzeri к факторам внешней среды и различным биоцидам проявляется на том же уровне, что и у других представителей рода. Однако характерной особенностью Р. stutzeri является их способность к деградации различных поллютантов (отходов производства), оставаясь резистентными к ним. Так, штамм может быть резистентным к таким веществам, как трибутилин, пестицид β-цифлутрин. Отмечена устойчивость Р. stutzeri к налидиксовой кислоте, хлорамфениколу, сульфаметоксазолу, n-гексану и др.
г) Антигенные свойства Р. stutzeri. Белки клеток Р. stutzeri, изученные в электрофорезе в полиакриламидном геле оказались очень специфичными для штаммов. Они могут использоваться при типировании и классификации микроорганизма. Свойства белков находились в соответствии с геномоварами культур. Такой результат не удивителен, потому что эти белки кодируются генами всего генома, а геномовары определяют по скрещиванию тотальной ДНК-ДНК.
Липополисахарид (ЛПС) является главной антигенной молекулой на клеточной поверхности. Считают, что это термостабильный О-антиген рода Pseudomonas. Специфичность антител связана с составом полисахаридных цепей внешней поверхности клетки. Среди семи различных геномоваров, с которыми были проведены эксперименты, выявлено серологическое разнообразие. Известно также, что взаимосвязь имеется только у близкородственных штаммов одного и того же геномовара. Белки внешней мембраны при анализе с додецелесульфатом натрия или в иммуноблоттинге с использованием поликлональных антисывороток к целым клеткам показывают аналогичный результат для всех штаммов, независимо от принадлежности к геномовару.
д) Патогенность. После 1956 г. в течение 15-летнего периода появилось несколько сообщений о выделении Р. stutzeri из клинического и патологического материала. Однако не было четкой взаимосвязи между этими культурами и инфекционным процессом. Так, 15 из 17 штаммов, изученных в 1966 г. R. Y. Stanier, имели клиническое происхождение. В 1973 г. в литературе описан первый хорошо документированный случай инфекции, вызванной Р. stutzeri после перелома голени. Затем появилось несколько сообщений о случаях инфицирования Р. stutzeri, сопровождавшихся бактериальной септицемией, об инфекции костной ткани после перелома, остеомиелите, а также при артрите, эндокардите, инфекции глаз (эндофтальмит и панофтальмит), менингите, пневмонии и эмпиеме, кожной инфекции, например, гангренозной эктиме, инфекциях уринарного тракта и вентрикулите.
При этих инфекциях только два случая оказались летальными, что указывает на относительно низкую вирулентность Р. stutzeri. Однако имеются сомнения, что смерть и в этих 2-х случаях была вызвана Р. stutzeri, т. к. у одного хронического больного наступила почечная недостаточность, а другой имел хроническую болезнь печени. Следует также заметить, что почти все пациенты относились к группе риска: имели основное заболевание, хирургическое вмешательство с последующим внутрибольничным заражением, предыдущую травму или кожную инфекцию, иммунодефициты. В двух случаях фактором риска был остеомиелит позвоночника, а у одного четырехлетнего мальчика — пневмония и эмпиема.
С целью определения степени распространения Р. stutzeri в госпиталях были проведены соответствующие исследования в двух различных университетских больницах в течение определенного периода. Объектами изучения являлись образцы гноя из раны, кровь, моча, трахейный аспират, слюна. Результаты изучения показали, что Р. stutzeri были выделены в 1-2 % из всех случаев инфекций, вызванных Pseudomonas. Сопоставимые данные (1,8%) получены от пациентов, имевших псевдомонадную инфекцию в сочетании с вирусом иммунодефицита. Наивысший уровень выделения Р. stutzeri имел место для образцов мочи.
Следовательно, можно заключить, что Р. stutzeri является убиквитарным микроорганизмом госпитальной среды. Вместе с тем, по-видимому, это редкий условно-патогенный микроорганизм.
- Читать далее "Методы диагностики инфекции Pseudomonas stutzeri и его выделение"
Редактор: Искандер Милевски. Дата публикации: 8.4.2020
- Пути распространения синегнойной палочки (Pseudomonas aeruginosa)
- Бактериологическая диагностика синегнойной палочки (P. aeruginosa)
- Определение чувствительности синегнойной палочки к антибактериальным препаратам (методы определения и критерии оценки)
- Лечение и профилактика инфекции синегнойной палочки (P. aeruginosa)
- Pseudomonas stutzeri: морфология, культуральные, биохимические свойства
- Методы диагностики инфекции Pseudomonas stutzeri и его выделение
- Флуоресцирующая псевдомонада (Pseudomonas fluorescens): морфология, культуральные, биохимические свойства
- Прочие псевдомонады: Pseudomonas putida, Pseudomonas alcaligenes, Pseudomonas diminuta, Pseudomonas mendocina, Pseudomonas pertucinogena, Pseudomonas pseudoalcaligenes, Pseudomonas vesicularis