Гранулоцитопоэз. Механизмы формирования нейтрофилов и макрофагов
Процесс, в результате которого гемопоэз переходит из желточного мешка в печень и затем в костный мозг, заключается не просто в изменении локализации, потому что каждая анатомическая структура продуцирует уникальные для нее виды клеток. Например, желточный мешок человеческого эмбриона не продуцирует нейтрофилы. Печень человеческого плода продуцирует нейтрофилы в очень малом количестве если вообще их продуцирует.
Эти немногочисленные нейтрофилы, обнаруживаемые в печени плода не организованы в гемопоэтические островки, как развивающиеся печеночные эритробласты, а располагаются главным образом около сосудов на значительном расстоянии друг от друга, что позволяет предположить, что они были образованы в другом месте и занесены в печень кровотоком. Нейтрофилы не образуются и в селезенке человеческого плода. В селезенке обнаруживаются в основном зрелые, равномерно распределенные нейтрофилы, что позволяет предположить, что они, так же как и нейтрофилы в печени, были образованы в другом месте и занесены сюда кровотоком.
Нейтрофилы впервые обнаруживаются у плода человека на 5-й неделе после оплодотворения в виде маленьких скоплений (кластеров) клеток вокруг аорты. Эти клетки содержат миелопероксидазу и созревают в клетки с сегментными ядрами; о других аналогиях между ними и нейтрофилами взрослых пока неизвестно. Пространство для костного мозга начинает формироваться примерно на 8-й неделе внутриутробного развития. Начиная с 8-10-й недели пространство костного мозга увеличивается, но нейтрофилы обнаруживаются в этом пространстве не ранее 10,5 нед. Эти первые нейтрофилы костного мозга имеют круглые ядра, содержат миелопероксидазу и имеют поверхность, характерную для миелобластов и промиелоцитов.
Начиная с 14-й недели и до конца внутриутробного развития наиболее распространенным типом клетки в пространстве костного мозга является нейтрофил.
У детей и взрослых нейтрофилы и макрофаги образуются из общего предшественника. Пока не выяснено, происходит ли это аналогичным образом у плода, потому что нейтрофилы и макрофаги появляются у него в разное время и в разных анатомических точках. До того как сформируется полость костного мозга, макрофаги вначале появляются в желточном мешке, печени, легких и головном мозге.
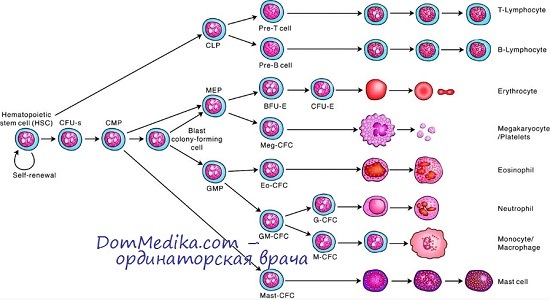
Механизмы, регулирующие образование гранулоцитов у плода, пока еще точно не установлены. Вполне вероятно, что, как и у взрослых, в этом процессе участвует гранулоцитарный колониестимулирующий фактор (Г-КСФ) и макрофагальный колониестимулирующий фактор (М-КСФ), так как они экспрессируются в развивающемся костном мозге плода уже на 6-й неделе после оплодотворения, в печени — на 8-й неделе. В тканях плода также широко распространены гранулоцитарный макрофагальный колониестимулирующий фактор (ГМ-КСФ) и фактор стволовой клетки (ФСК).
Тем не менее пока не выявлено никаких изменений экспрессии мРНК какого-либо из этих факторов или их специфических рецепторов, а также изменений в концентрации белков (что определяется иммуногистохимическим окрашиванием), которые могли бы служить сигналом для образования у плода нейтрофилов и макрофагов. Точные сигналы пока не идентифицированы.
До III триместра в кровотоке обнаруживается мало нейтрофилов. Форестер (Forester) и Даффос (Daffos) определили, что среднее количество циркулирующих нейтрофилов у плодов на 20-й неделе гестации составляет 190/мм3 (диапазон — от 0 до 490/мм3). Хотя кровь плода содержит очень мало зрелых нейтрофилов, в ней относительно много клеток-предшественниц, способных генерировать клоны нейтрофилов. При культивировании этих клеток-предшественниц плода in vitro в присутствии рекомбинантного Г-КСФ они созревают в большие колонии нейтрофилов.
Физиологическая роль Г-КСФ заключается в регулировании образования нейтрофилов в сторону увеличения; по-видимому, они выполняют эту роль и у плода, и у новорожденного, и у взрослого человека. Таким образом, малые количества циркулирующих и резервных нейтрофилов во II триместре могут быть частично обусловлены продуцированием малого количества Г-КСФ. Моноциты, выделенные из крови взрослого человека, продуцируют этот фактор при стимулировании различными медиаторами воспаления, например липополисахаридами бактерий или ИЛ-1. В отличие от этого, моноциты, выделенные из пуповинной крови недоношенных детей и из печени и костного мозга выкидышей на сроке до 24 нед., продуцируют лишь небольшое количество (в 10-100 раз меньше на клетку) белка Г-КСФ и мРНК при стимулировании липополисахаридами бактерий или ИЛ-1. Несмотря на такую низкую способность продуцировать Г-КСФ, нейтрофилы новорожденных несут на своей поверхности такое же количество рецепторов Г-КСФ, с такой же аффинностью, что и нейтрофилы взрослого.
Действие Г-КСФ, М-КСФ, ГМ-КСФ и ФСК у плода не ограничивается гемопоэзом. Рецепторы для каждого из этих факторов находятся в четко определенных участках ЦНС и ЖКТ плода, и характер их экспрессии меняется по мере развития. Помимо известного участия в гемопоэзе эти факторы играют другие важные роли в развитии плода.
- Читать "Тромбоцитопоэз - механизмы формирования тромбоцитов. Тромбопоэтин"
Оглавление темы "Анемии у детей":- Формирование гемопоэза. Гемопоэз плода - эмбриона
- Гранулоцитопоэз. Механизмы формирования нейтрофилов и макрофагов
- Тромбоцитопоэз - механизмы формирования тромбоцитов. Тромбопоэтин
- Эмбриональный и фетальный гемоглобины
- Соотношение разных видов гемоглобинов в организме. Гемоглобин при болезнях
- Метаболизм и время жизни эритроцитов
- Анемия у детей. Причины
- Транзиторная эритробластопения детей (ТЭД). Красноклеточные аплазии
- Анемии при заболеваниях почек у детей. Диагностика и лечение
- Врожденные дизэритропоэтические анемии (ВДА) у детей. Диагностика и лечение