Морфология и культуральные свойства хеликобактер пилори (H. pylori)
Форма отдельных клеток Н. pylori в значительной степени зависит от условий его существования. В комфортных условиях (в желудке при нелеченных хроническом гастрите и язвенной болезни либо при культивировании in vitro в оптимальном режиме) он имеет строение, расценивающееся как типичное. Различают два типичных морфологических варианта Н. pylori — почковидный и изогнутый палочковидный. Первый представляет собой слегка и строго посредине изгибающиеся палочковидные клетки длиной от 1,5 до 3,0 мкм и диаметром от 0,3 до 0,5 мкм. Поскольку длина таких клеток не слишком превосходит их ширину, они приобретают очертания, напоминающие человеческую почку.
Второй морфологический вариант — несколько более крупные палочковидные клетки длиной от 2,0 до 6,5 мкм и диаметром от 0,5 до 0,8 мкм, плавно S-образно изогнутые по плоскости вокруг длинной оси клетки, причем точка изгиба, как правило, располагается ближе к одному из ее полюсов. Зачастую две клетки не расходятся после деления, оставаясь сцепленными своими полюсами. В подобных случаях две изогнутые в несовпадающих плоскостях бактериальные клетки образуют извитую в пространстве биклеточную структуру.
Н. pylori — грамотрицательная бактерия, весьма слабо окрашивающаяся анилиновыми красителями. Несравненно более эффективно серебрение, позволяющее получать воспроизводимые результаты вне зависимости от возраста исследуемой культуры.
Н. pylori является лофотрихом; один из полусферических полюсов его клеток снабжен пучком жгутиков, число которых варьирует от четырех до семи. Длина жгутика, составляющая 5-10 мкм, как видно, несколько превосходит длину несущей его бактериальной клетки; диаметр жгутика — 30 нм.
Клеткам Н. pylori в полной мере свойственна ультраструктура прокариотических клеток грамнегативного типа. Их цитоплазма, окруженная цитоплазматической мембраной и не содержащая каких-либо мембранных органелл, характеризуется наличием электронно-плотного рибонуклеопротеидного комплекса, образованного скоплениями рибосом и центрально расположенным нуклеоидом. Снаружи цитоплазматическая мембрана окружена клеточной стенкой, которая состоит из тонкого регулярного слоя высокоплотного для электронов пептидогликана (так называемого ригидного слоя) и покрывающей его трехслойной наружной мембраны; прозрачное для электронов пространство между цитоплазматической и наружной мембранами обозначается как периплазматическое.
В естественных условиях существования, но не при культивировании in vitro, клетки Н. pylori покрыты микрокапсулой.
Описанная «классическая» ультраструктура характерна лишь для почковидных вариантов клеток Н. pylori, а уже изогнутые палочковидные клетки имеют некоторые дефекты клеточной стенки (небольшое уплотнение наружной мембраны, незначительное нарушение регулярности строения ригидного слоя, едва выраженное расширение периплазматического пространства), являющиеся ультраструктурными маркерами старения бактериальных клеток. В бактериальных клетках биклеточных структур эти изменения более выражены.
Морфологические изменения клеток наиболее четко прослеживаются при культивировании Н. pylori на искусственных питательных средах. Так, в ранней логарифмической фазе роста, когда культура пребывает в достаточно комфортных для нее условиях, она представлена почти исключительно почковидными клетками, лишенными каких-либо ультраструктурных дефектов. В поздней логарифмической фазе роста в популяции отмечается появление палочковидных клеток, которые к началу стационарной фазы полностью вытесняют почковидные. По мере истощения питательных субстратов и накопления метаболитов все более выраженными становятся дефекты клеточной стенки: во все большей степени нарушается структура ригидного слоя вплоть до возникновения его дефектов. Кроме того, отмечается прогрессирующая деструкция рибонуклеопротеидного комплекса.
Следствием нарушения структуры ригидного слоя является возникновение бактериальных клеток неправильной (отклоняющейся от палочковидной) формы. Более того, по мере нарастания дефектов в культуре возрастает число бактериальных клеток сферопластного и протопластного типов и соответственно форм, имеющих дефектную клеточную стенку и вовсе лишенных таковой. Понятно, что пептидогликан с обширными дефектами оказывается не в состоянии выполнять функцию полноценного наружного скелета бактериальной клетки (т.е. обеспечивать ей сохранение изогнутой палочковидной формы). По этой причине сферопласты и тем более вовсе лишенные пептидогликана протопласты принимают наиболее выгодную энергетически сферическую форму.
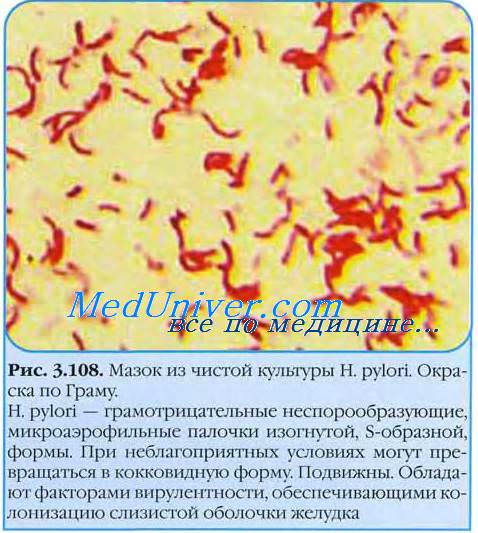
В естественных условиях существования популяцию Н. pylori составляют бактериальные клетки всех описанных выше типов. При этом может быть отмечено преобладание того или иного морфологического варианта в различных отделах желудка. В ходе проведения иротивохеликобактерной эрадикационной терапии в популяции Н. pylori явно преобладают формы с выраженными дефектами клеточной стенки — сферопласты и протопласты.
Способность клеток Н. pylori трансформироваться в округлые по форме — сферопласты и протопласты послужила причиной многих заблуждений. Наиболее значительным является введение в гастроэнтерологическую лексику термина «кокковидные формы Н. pylori» — в корне неверного и подменяющего собой термины «сферопласт» и «протопласт».
Как видно из вышесказанного, Н. pylori свойственна выраженная ультраструктурная и морфологическая лабильность — способность в зависимости от условий существования легко изменять свою клеточную структуру и форму.
Благодаря наличию лофотрихиально расположенного пучка жгутиков клетки Н. pylori способны к активной локомоции — вращательно-поступательному (штопорообразному) движению безжгутиковым полюсом вперед. При этом отдельные клетки достигают изрядной скорости перемещения — 2-3 мкм/с в воде и 0,5-1 мкм/с в водном растворе глицерина, вязкость которого сопоставима с вязкостью желудочной слизи.
Жгутиковый мотор функционирует по принципу протоновой помпы: создает направленное перемещение протонов внутрь клетки, сопровождающееся освобождением энергии. Освобожденная энергия расходуется на вращение жгутика без каких-либо существенных потерь, что дает основание рассматривать жгутик как один из наиболее эффективных биологических механизмов.
Движение клеток Н. pylori носит отнюдь не беспорядочный характер; им свойствен таксис различного типа — направленное перемещение против градиента концентрации привлекающих их химических соединений. К числу последних относятся мочевина, бикарбонаты, аминокислоты — аттрактанты, в изобилии присутствующие на наружной стороне пласта поверхностного эпителия желудка. Каждому из аттрактантов соответствует белковый рецептор Н. pylori, интегрированный в цитоплазматическую мембрану и выступающий в периплазматическое пространство. Результатом взаимодействия рецептора с аттрактантом является сигнал, активирующий работу жгутикового мотора и, как следствие, ускорение движения бактериальной клетки. Помимо перечисленных выше низкомолекулярных аттрактантов, обеспечивающих положительный хемотаксис Н. pylori, в роли аттрактанта выступает и высокомолекулярный муцин — важнейший структурный белок желудочной слизи, обусловливающий его положительный мукотаксис.
Наконец, Н. pylori свойствен положительный вискотаксис: весьма редкий вид таксиса в направлении возрастания вязкости — физического фактора, характерного для жидких сред. Вязкость желудочной слизи, как и концентрация в ней химических соединений различной природы, возрастает в направлении «просвет желудка — эпителиальный пласт», достигая максимума на поверхности последнего.
Сферопласты и тем более протопласты, по-видимому, лишены жгутикового аппарата и способны лишь к хаотическому броуновскому движению, вряд ли имеющему какое-либо физиологическое значение.
Таким образом, способность Н. pylori к активной локомоции, регулируемой факторами различной природы, обеспечивает ему в естественных условиях существования эффективную возможность достижения слизистой оболочки желудка.
Энзиматический профиль культур Н. pylori достаточно широк, его формируют ферменты различных классов, локализующиеся как в цитозоле бактериальной клетки, так и в толще цитоплазматической и наружной мембран. Зачастую ферменты Н. pylori выполняют двойственную функцию: участвуют в метаболизме и оказывают повреждающее действие на слизистую оболочку желудка, иными словами, выступают в качестве факторов патогенности.
Для Н. pylori характерно наличие оксидоредуктаз (оксидаз) нескольких типов, локализующихся преимущественно в цитоплазматической мембране и принимающих участие в дыхательном метаболизме, катализируя окисление восстановленных форм, а также превра!цение пирувата в ацетилкоэнзим А — ключевой метаболит синтеза пептидогликана, фосфолипидов, жирных кислот, ароматических аминокислот и др.
Н. pylori обладает высокой каталазной активностью — продуцирует каталазу, катализирующую образование воды из перекиси водорода, лишая тем самым последнюю способности к генерации пероксид-иона O22- — токсичной для живой материи формы кислорода. Другую токсичную форму кислорода — супероксид-ион O22- нейтрализует поверхностно локализующаяся супероксид-дисмутаза — второй антиоксидантный фермент Н. pylori.
Кроме того, Н. pylori синтезирует дегидрогеназы нескольких типов, участвующие в дыхательном метаболизме. Отдельного упоминания заслуживает алкоголь-дегидрогеназа, катализирующая восстановление ацетальдегида до этанола на конечной стадии ферментации глюкозы. Однако при высоких концентрациях как экзо-, так и эндогенного этанола этот фермент способен катализировать обратный процесс — окисление этанола до ацетальдегида.
Н. pylori свойственно наличие семейства протеаз, среди которых особого внимания заслуживает сульфогидролаза муциназа — фермент, осуществляющий гидролиз богатого сульфидами муцина.
Наконец, Н.pylori способен продуцировать фосфолипазы нескольких типов, катализирующие гидролиз фосфолипидов цитоплазматических мембран эукариотических клеток.
Помимо перечисленных выше ферментов, в культурах Н. pylori обнаруживаются кислые и гцелочные фосфатазы, АТФазы, экзо- и эндонуклеазы, однако наиболее заметное положение, безусловно, занимает уреаза — амидогидролаза мочевины. По уровню продукции этого весьма распространенного в природе фермента Н. pylori не имеет себе равных ни среди прокариотических, ни среди эукариотических организмов. Уреаза является характеристическим для вида Н. pylori ферментом и мажорным белком его культур, на ее долю приходится более 5% их общего белка.
Транспорт воды и ионов осуществляется путем простой диффузии, низкомолекулярных водорастворимых метаболитов — путем облегченной диффузии, высокомолекулярные же метаболиты транспортируются посредством активного транспорта, осуществляющегося через многочисленные поры наружной мембраны — каналы, сформированные особыми белками — поринами, и соединяющие периплазматическое пространство с внешней средой.
Н. pylori использует глюкозу в качестве единственного источника углеводов и одного из важнейших источников энергии. В качестве прочих источников энергии выступают серин, аланин, аспарагин и пролин. Источником экзогенного азота, необходимого Н. pylori, в частности для синтеза нуклеотидов, являются аммоний, аланин, серин и глутамин.
Будучи микроаэробным микроорганизмом, Н. pylori в зависимости от условий окружающей среды избирает аэробный либо анаэробный пути дыхания, принципиально различными путями метаболизируя глюкозу и пируват: при помощи альдолаз — в первом и дегидрогеназ — во втором случае. Поскольку Н. pylori не способен к осуществлению замкнутого цикла трикарбоновых кислот, чрезвычайно важную для его анаэробного дыхания роль играет фумаратредуктаза, сочетающая цикл Кребса с иными метаболическими путями.
Таким образом, Н. pylori имеет весьма сложный биохимический состав и своеобразный метаболизм, складывающийся из оптимального для конкретных условий окружающей среды сочетания аэробного и анаэробного путей освобождения энергии. При культивировании in vitro оптимальное сочетание этих путей складывается, повидимому, в атмосфере со сниженным содержанием кислорода и повышенным — углекислоты — в микроаэробной капнофильной атмосфере.
- Читать далее "Устойчивость хеликобактер пилори (H. pylori) к факторам внешней среды"
Редактор: Искандер Милевски. Дата публикации: 9.1.2020
- Хеликобактер (Helicobacter pylori): таксономия
- Морфология и культуральные свойства хеликобактер пилори (H. pylori)
- Устойчивость хеликобактер пилори (H. pylori) к факторам внешней среды
- Антигены и генетика хеликобактер пилори (H. pylori)
- Факторы патогенности хеликобактер пилори (H. pylori)
- Патогенез инфекции хеликобактер пилори (H. pylori)
- Эпидемиология хеликобактер пилори (H. pylori)
- Микроскопический метод диагностики хеликобактер пилори (H. pylori)
- Бактериологический метод диагностики хеликобактер пилори (H. pylori)
- Методы экспресс-диагностики хеликобактер пилори (H. pylori)
- Серологическая диагностика хеликобактер пилори (H. pylori)
- Лечение и профилактика инфекции хеликобактер пилори (H. pylori)